DIVISION OF SPECIATION MECHANISMS II
1) from Purdue University, West Lafayette, IN, USA
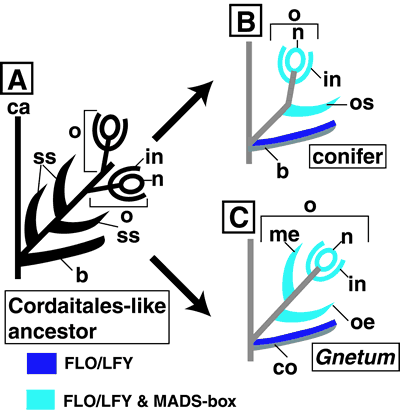
All living organisms evolved from a common ancestor more than 3.5 billion years ago, and accumulated mutations on their genomes caused the present biodiversity. The traces of evolutionary processes are found in the genomes of extant organisms. By comparing the genomes of different organisms, we can infer (1) the phylogenetic relationships of extant organisms and (2) the genetic changes having caused the evolution of morphology and development. The inferred phylogenetic relationships give important insights on problems in various fields of evolutionary biology, and our group focuses on biogeography, evolution of morphological traits, and systematics in wide range of taxa. On the evolution of morphology and development, we aim to explore genetic changes led the evolution of plant body plan. We selected Arabidopsis (angiosperm), Gnetum (gymnosperm), Ginkgo (gymnosperm), Ceratopteris (pteridophyte), Physcomitrella (bryophyte), and some green algae as models to compare the gene functions involved in development of the reproductive organs and shoot apical meristem of land plants. I. Evolution of reproductive organs in land plantsA flower is the most complex reproductive organ in land plants and composed of sepals, petals, stamens, and gynoecium. Female haploid reproductive cells are covered with a sporangium (nucellus) and two integuments, and further enclosed in a gynoecium. Male haploid reproductive cells (pollens) are covered with a sporangium (pollen sack). On the other hand, gymnosperms and ferns have simpler reproductive organs than angiosperms and lack sepals and petals. Female sporangia (nucellus) of gymnosperms are covered with only one integument. Sporangia of ferns have no integuments and are naked on the abaxial side of a leaf. The development of floral organs is mainly regulated by A-, B-, C-function genes, which are members of the MADS-box gene family. These genes are transcription factors containing the conserved MADS and K domains. MADS-box genes of angiosperms are divided into more than 10 groups based on the gene tree. The LEAFY gene is the positive regulator of the MADS-box genes in flower primordia. What kind of changes of the MADS-box genes caused the evolution of the complex reproductive organs in the flowering plant lineage ? Comparisons of MADS-box and LFY genes in vascular plants suggest that the following sequential changes occurred in the evolution of reproductive organs. (1) Plant-type MADS-box genes with both MADS and K domains were established. (2) The number of MADS-box genes increased, and the three ancestral MADS-box genes that later generate A-, B-, C-functions genes were likely originated before the divergence of ferns and seed plants. (3) Specifically expressed MADS-box genes in reproductive organs evolved from generally expressed ones in the seed plant lineage. (4) The ancestral gene of the AG group of MADS-box genes acquired the C-function before the divergence of extant gymnosperms and angiosperms. (5) The gene duplication that formed the AP3 and PI groups in MADS-box genes occurred before the diversification of extant gymnosperms and angiosperms. (6) The ancestral gene of angiosperm A-function gene was lost in extant gymnosperm lineage. (7) LFY gene became positively regulate MADS-box genes after the divergence of ferns and seed plants, because the fern LFYgene does not directly regulate MADS-box genes (Himi et al. 2001). The FLO/LFYgene phylogenetic tree indicates that both duplication and loss of FLO/LFYhomologs occurred during the course of vascular plant evolution. The inductive pathway from the LFYgene to the MADS-box genes already existed in the common ancestor of angiosperms and gymnosperms, because overexpression of GnetumLFY homolog in transgenic Arabidopsispromoted a conversion of a shoot meristem to a floral primordium and the Arabidopsis LFYnull mutant, lfy-26,with a malformed flower, was complemented by overexpression of GnetumLFYgene. (8) Spatial and temporal patterns of A-, B-, C-function gene expression were established in the angiosperm lineage (Shindo et al. 2001). II. Homology of reproductive organs in seed plantsThe morphological variation among reproductive organs of extant seed plants makes assessment of organ homology difficult. Comparisons of expression patterns of homeotic genes that control organ development will yield new information about the homology of organs to assess inferences deduced from previous morphological studies. To provide insights into the evolution of reproductive organs in seed plants, a Gnetum LFYhomolog was cloned and its expression patters were compared to the conifer LFYhomolog. The comparison suggests that the Gnetumcollar and ovule are homologous with the conifer bract and ovule-ovuliferous scale complex, respectively. This inference is concordant with our previous comparisons of expression patters of orthologous MADS-box genes between Gnetum and conifers (Shindo et al. 2001).
III-I. Evolution of vegetative organsThe ancestor of land plants was primarily haploid. The only diploid cell was the zygote, which immediately underwent meiosis. It is believed that early during land plant evolution, zygotic meiosis was delayed and a multicellular diploid sporophytic generation became interpolated into the life cycle. In the early stages of land plant evolution, sporophytes are epiphytic to gametophytes, as observed in extant bryophytes. During the course of evolution, both generations started to grow independently at the stage of pteridophytes. Finally gametophytes became much reduced and epiphytic to sporophytes in seed plant lineage. Molecular mechanisms of development in a diploid generation have been well studied in some model angiosperms, but we have scarce information on those in a gametophyte generation. For example, mosses have leaf- and stem-like organs in their haploid generation, but it is completely unknown whether similar genes involved in angiosperm leaf and stem development are used in the gametophytic generation of mosses or not. To understand the evolution of body plans in diploid and haploid generation at the molecular level, we focus on the comparison of molecular mechanisms governing shoot development between Arabidopsis and the moss Physcomitrella patens. P. patens is known by its high rate of homologous recombination and suitable for analyze gene functions using the gene targeting, and should be a good model lower land plants. III-II. Characterization of homeobox genes in the moss Physcomitrella patensHomeobox genes encode transcription factors involved in many aspects of developmental processes including shoot development in angiosperms. The homeodomain-leucine zipper (HD-Zip) genes, which are characterized by the presence of both a homeodomain and a leucine zipper motif, form a clade within the homeobox superfamily and previously reported only from vascular plants. We isolated 10 HD-Zip genes from P. patens(Pphb1-10genes). Based on a phylogenetic analysis of the 10 Pphbgenes and previously reported vascular plant HD-Zip genes, all the Pphb genes except Pphb3 belong to three of the four HD-Zip subfamilies (HD-Zip I, II, and III), indicating that these subfamilies originated before the divergence of the vascular plant and moss lineages. Pphb3is sister to HD-Zip II subfamily, and has some distinctive characteristics, including the difference of a1and d1sites of its leucine zipper motif, which are well conserved in each HD-Zip subfamily. Comparison of the genetic divergence of representative HD-Zip I and II genes showed that the evolutionary rate of HD-Zip I genes was faster than HD-Zip II genes (Sakakibara et al. 2001). The moss homologs of SHOOTMERISTEMLESS and ZWILLE genes, which are involved in Arabidopsis shoot development, have been cloned and their characterization is in progress. III-III. Establishment of enhancer and gene trap lines in the moss Physcomitrella patensWe also established enhancer and gene trap lines and tagged mutant libraries of P. patens to clone genes involved in the leafy shoot development in haploid generation (Hiwatashi et al. 2001). Elements for gene-trap and enhancer-trap systems were constructed using the uidA reporter gene with either a splice acceptor or a minimal promoter, respectively. Through a high rate of transformation conferred by a method utilizing homologous recombination, 235 gene-trap and 1073 enhancer-trap lines were obtained from 5637 and 3726 transgenic lines, respectively. Expression patterns of these trap lines in the moss gametophyte varied. The candidate gene trapped in a gene-trap line YH209, which shows rhizoid-specific expression, was obtained by 5' and 3' RACE. This gene was named PpGLU, and forms a clade with plant acidic alpha-glucosidase genes. Thus, these gene-trap and enhancer-trap systems should prove useful to identify tissue- and cell-specific genes in Physcomitrella.
Publication List:Fujita, T., Ikeda, M., Kusano, S., Yamazaki, S., Ito, S., Obayashi, M., Yanagi, K. (2001) Amino acid substitution analyses of the DNA contact region, two amphipathic a-helices and a recognition-helix-like helix outside the dimeric b-barrel of Epstein-Barr virus nuclear antigen 1. Intervirology. 44: 271-282. Himi, S., Sano, R., Nishiyama, T., Tanahashi, T., Kato, M., Ueda, K. and Hasebe, M. (2001) Evolution of MADS-box gene induced by FLO/LFYgenes. J. Mol. Evol.53: 387-393. Hiwatashi, Y., Nishiyama, T., Fujita, T. and Hasebe, M. (2001) Establishment of gene-trap and enhancer-trap systems in the moss Physcomitrella patens. Plant J.28: 1-14. Kurumatani, M., Yagi, K., Murata, T., Tezuka, M., Mander, L. N., Nishiyama, M. and Yamane, H. (2001) Isolation and identification of antheridiogens in the ferns, Lygodium microphyllum and Lygodium reticulatum. Biosci. Biotechnol. Biochem. 65: 2311-2314. Sakakibara, K., Nishiyama, T., Kato, M. and Hasebe, M. (2001) Isolation of Homeodomain-Leucine Zipper Genes from the Moss Physcomitrella patens and the Evolution of Homeodomain-Leucine Zipper Genes in Land Plants. Mol. Biol. Evol. 18: 491-502. Shindo, S., Sakakibara, K., Sano, R., Ueda, K. and Hasebe, M. (2001) Characterizatin of a FLORICAULA/LEAFYhomologue of Gnetum parvifolium, and its implications for the evolution of reproductive organs in seed plants. Int. J. Plant Sci.162: 1199-1209. Yoshihara, S., Geng, X. X., Okamoto, S., Yura, K., Murata, T., Go, M., Ohmori, M. and Ikeuchi, M. (2001) Mutational analysis of genes involved in pilus structure, motility and transformation competency in the unicellular motile cyanobacterium Synechocystissp. PCC 6803. Plant Cell Physiol. 42: 63-73. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |