DIVISION OF MORPHOGENESIS
* from September 1, 2001
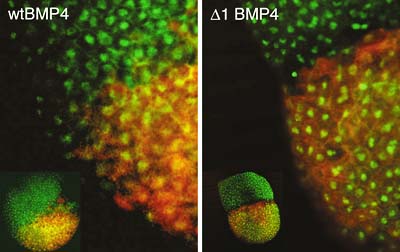
The complex morphogenesis of organisms is achieved by consecutive cell-to-cell interactions during development. Recent studies suggest that growth factors play crucial roles in controlling such intercellular communications in a variety of organisms. In addition to secretory factors, transcription factors which act cell-autonomously are thought to be essential for the determination of cell fates. Our main interest is to know how pattern formation and morphogenesis during development is regulated by these growth factors and transcription factors. We address this problem using several model animals, including frog, fly, acidian and nematode, employing embryology, genetics, molecular and cellular biology, and biochemistry. In addition, we have recently introduced array technology to elucidate precise genetic program controlling early development. I.Regulation of growth factor diffusion in pattern formationDuring early development, cells receive positional information from neighboring cells to form tissue patterns in initially uniform germ layers. Ligands of the transforming growth factor (TGF-b) superfamily are known to participate in this pattern formation. In particular, activin has been shown to act as a long-range dorsalizing signal to establish a concentration gradient in Xenopus. In contrast, the action of BMP-2 and BMP-4, another members of the family, appear to influence and induce ventral fates only where they are expressed. This raises a question as to how the action of BMPs is tightly restricted within and around the cells that produce them. We have demonstrated that a basic core of only three amino acids in the N-terminal region of BMP-4 is required for its restriction to the non-neural ectoderm corresponding to its expression domain. Our results also suggest that heparan sulfate proteoglycans bind to this basic core and thus play a role in trapping BMP-4. We propose that restriction of BMP diffusion in vivo through the interaction with extracellular environment is critical for the precise definition of non-neural ectoderm during early embryogenesis. II. Gastrulation movement regulated by Wnt signalingGastrulation is one of the most important processes during morphogenesis of early embryo, involving dynamic cell migration and change in embryo shape. Almost all animals undergo gastrulation to form the gut. In spite of its importance, the mechanism underlying the event has just begun to be studied at molecular level. The Wnt family of secretory glycoprotein is one of the major families of developmentally important signaling molecules and plays important roles in embryonic induction, generation of cell polarity, and specification of cell fate. Wnt proteins bind Frizzled (Fz) receptors, a membrane protein with seven transmembrane domains. So far, the Wnt/Fz signaling cascade has been shown to branch into at least three pathways. These are the canonical pathway, the planar cell polarity (PCP) pathway, and the pathway through protein kinase C (PKC). The canonical pathway is the best-characterized signaling cascade which involves Dishevelled (Dsh/Dvl) and b-catenin and contributes to establish the dorsoventral axis in Xenopusembryos. The PCP pathway is essential for cell polarization and cell movement during gastrulation. This pathway seems to involve Dsh/Dvl and c-Jun N-terminal kinase (JNK). Although these signaling pathways have been investigated extensively, it is still unknown exactly how the Fz receptors generate signals to regulate gastrulation cell movement. Thus, we screened for proteins which interact with the cytoplasmic domains of Fz using the yeast two-hybrid assay. So far, we have cloned two Xenopus genes. One encodes a protein which contains two PDZ domains. This gene is expressed ubiquitously during gastrulation, and later in the central nervous system. Our preliminary results suggest its role in the Fz signaling, particularly in the PCP pathway. (a) It binds Fz-7, an important receptor in the PCP signaling pathway in Xenopusembryos, more strongly than other Fzs. (b) Overexpression of a predicted dominant-negative mutant, causes severe gastrulation defect, a typical phenotype seen by the inhibition of the PCP pathway. These results suggest that this protein may be an essential component for gastrulation cell movement acting in the PCP pathway. III. Genetic screening for novel DPP/BMP signaling components utilizing Drosophila model systemDrosophila is one of the ideal model organisms to dissect signal transduction pathway by genetic methods. We have carried out dominant suppressor screening for two transgenic mutant flies expressed constitutively activated DPP/BMP type-I receptors in wing imaginal discs. We isolated 19 suppressor mutants, Suppressor of constitutively activated Dpp singnaling(Scad). Alleles of punt, Mad, shn and dCrebA were found in isolated Scad mutants. We now focus to study a novel locus Scad67. Scad67 encodes a protein with a SP-RING motif. SP-RING motif was originally found in PIAS- family proteins including Drosophila PIAS homolog Su(var)2-10/Zimp. Recent studies have shown that PIAS-family proteins function as a E3-SUMO ligase and enhance SUMO conjugation against specific substrates. Scad67is an evolutionary conserved molecule and we found two Scad67homologous genes in human and also in other vertebrates. Homozygous Scad67 mutants show embryonic to pupal stage lethality. The most severe zygotic mutant shows embryonic head structure and segmentation defects. Detailed Scad67 function analysis especially in the SUMO mediated biological processes and also in DPP signalling are ongoing. We also have been interested in the in vivo function of the TGF-b activated kinase-1 (TAK1) in Drosophila. We succeed to isolate dTAK1null mutant in collaborated with B. Lemaitre's group (CNRS, France). Interestingly, dTAK1 mutation does shows morphological and viability defects. However, we observed an impairment of antibacterial peptide gene expression in dTAK1 mutants. Genetic studies of dTAK1 suggested an evolutionary conserved role for TAK1 in the control of rel/NF-kB dependent innate immune responses. IV. Brachyury downstream notochord differentiation in the ascidian embryoAscidians, urochordates, are one of the three chordate groups, and the ascidian tadpole is thought to represent the most simplified and primitive chordate body plan. It contains a notochord, which is a defining characteristic of chordate embryo composed of only 40 cells. To understand the morphogenesis in this simple system, we have focused on a gene, Brachyury, which is known to play an important role in the notochord development. In ascidian, Brachyury is expressed exclusively in the notochord and the misexpression of the Brachyury gene (Ci-Bra) of Ciona intestinalisis sufficient to transform endoderm into notochord. This gene encodes a sequence-specific activator that contains a T-box DNA-binding domain, and in vertebrates, it is initially expressed throughout the presumptive mesoderm and gradually restricted to the developing notochord and tailbud. The phenotype of the Brachyury mutants in mice and zebrafish revealed that this gene is essential for notochord differentiation. Our goal is to elucidate the down stream pathway of this important gene in ascidian in order to set the stage for understanding not only the formation and function of the notochord but how this important structure has evolved. We conducted the subtractive hybridization screens to identify potential Brachyury target genes that are induced upon Ci-Braoverexpression. Out of 501 independent cDNA clones that were induced cDNAs, 38 were specifically expressed in notochord cells (Fig. 2). We characterized 20 of them by determining the complete nucleotide sequences and in situ hybridization analyses which show the spatial and temporal expression patterns of the cDNAs. These potential Ci-Bra downstream genes appear to encode a broad spectrum of divergent proteins associated with notochord formation and function. V. TGF-b family in nematodeWe have previously shown that DBL-1, a member of TGF-b superfamily regulates body length in C. elegans. To understand molecular mechanism of body length regulation by DBL-1, we examined tissue-specific requirement of a DBL-1 receptor SMA-6 by rescuing sma-6mutation with tissue specific expression of the receptor. We found that hypodermal expression of SMA-6 is sufficient to rescue the sma phenotype, suggesting that hypodermis is the most critical target tissue of DBL-1 to regulate body length. Next we screened for target genes regulated by DBL-1 in hypodermal cells, and identified a gene yk298h6 whose disruption by dsRNAi resulted in long worm. yk298h6 was later found to be identical with lon-1, a gene encoding a type II membrane protein belonging to the PR family conserved from plant to human. LON-1 appears to suppress hypodermal polyploidization and thus negatively regulate body length.
Publication ListDong, W., Ogawa, S., Tsukiyama, S., Okuhara, S. Niiyama, M., Ueno, N., Peterson, R. E. and Hiraga, T.(2002) 2,3,7,8-Tetrachlorodibenzo-p-dioxin toxicity in the zebrafish embryo: altered regional blood flow and impaired lower jaw development. Teraoka, H., Toxicological Sciences65,192-199. Hanazawa, M., Mochii, M. Ueno, N., Kohara, Y. and Iino, Y. (2001) Use of cDNA subtraction and RNA interference screens in combination reveals genes required for germ-line development in Caenohabditis elegans. Proc. Natl. Acad. Sci. USA98, 8686-8691. Ishizuya-Oka, A., Ueda, S,, Amano, T., Shimizu, K., Suzuki, K., Ueno, N. and Yoshizato, K. (2001) Thyroid-hormone-dependent and fibroblast-specific expression of BMP-4 correlates with adult epithelial development during amphibian intestinal remodeling. Cell Tissue Res. 303, 187-195. Itoh, K., Udagawa, N., Katagiri, T., Iemura, S., Ueno, N., Yasuda, H., Higashio, K., Quinn, J. M., Gillespie, M. T., Martin, T. J., Suda, T., Takahashi, N. (2001) Bone morphogenetic protein 2 stimulates osteoclast differentiation and survival supported by receptor activator of nuclear factor-kappab ligand. Endocrinology.142, 3656-3662. Kurata T., Nakabayashi, J., Yamamoto, T.S., Mochii, M. and Ueno, N. (2001) Visualization of endogenous BMP signaling during Xenopus development. Differentiation67, 33-40. Mitani, Y., Takahashi, H. and Satoh, N. (2001) Regulation of the muscle-specific expression and function of an ascidian T-box gene, As-T2. Development128, 3717-3728. Morita, K., Shimizu, M., Shibuya, H. and Ueno, N. (2001) A Daf-1-binding protein BRA-1 is a negative regulator of Daf-7 TGF-b signaling. Proc. Natl. Acad. Sci. USA98, 6284-6288. Ohkawara, B., Iemura, S., ten Dijke, P. and Ueno, N. (2002) Action range of BMP is defined by its N-terminal basic amino acid core. Curr. Biol, 12, 205-209. Sakuta, H., Suzuki, R., Takahashi, H., Kato, A., Shintani, T., Iemura, S.-i, Yamamoto, T. S., Ueno, N. and Noda, M. (2001) Ventroptin: a BMP-4 antagonist expressed in a double-gradient pattern in the retina. Science293, 111-115. Sugawara, K. Morita, K., Ueno, N. and Shibuya, H. (2001) BIP, a BRAM-interacting protein involved in TGF-beta signalling, regulates body length in Caenorhabditis elegans. Genes Cells6, 599-606. Takahashi, H. and Satoh, N. (2001) Trunk lateral cell-specific genes of ascidian Halocynthia roretzi. Zool. Sci. 18, 361-366. Vidal, S. S., Khush, R. S., Leulier, F., Tzou, P., Nakamura, M. and Lemaitre, B. (2001) Mutations in the DrosophiladTAK1 gene reveal a conserved function for MAPKKKs in the control of rel/NF-kappaB-dependent innate immune responses. Genes. Dev.15, 1900-12. Yamamoto, T. S.,Takagi, C., Hyodo, A. C. and Ueno, N.(2001) Suppression of head formation by Xmsx-1through the inhibition of intracellualr nodal signaling. Development128, 2769-2779. Yoshida, S., Morita, K., Mochii, M. and Ueno, N. (2001) Hypodermal expression of Caenorhabditis elegansTGF-b type I receptor SMA-6 is essential for the growth and maintenance of body length. Dev. Biol.240, 32-45. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |