DIVISION OF CELL FUSION (ADJUNCT)
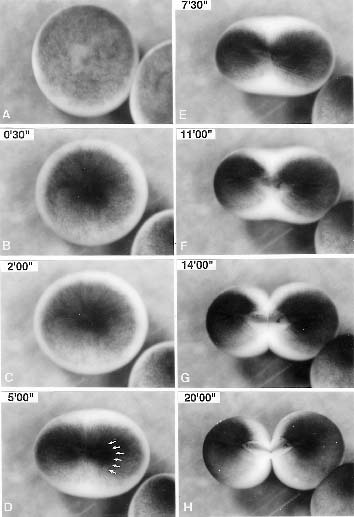
Cytokinesis in animal and some primitive eukaryotic cells is achieved by the progressive contraction of the cleavage furrow. The cleavage furrow contains a contractile apparatus, called the contractile ring, which is composed of a bundle of actin filaments that lies in the furrow cortex beneath the plasma membrane. It has been established that the contractile ring contracts as the result of interaction between actin filaments and myosin. However, little is known about process of its formation, mechanism that controls its formation, protein constituents, and its ultrastructure. The goal of our research is to solve these problems and thereby clarify the molecular mechanism of cytokinesis. For this purpose, we use three kinds of cells, namely, sea urchin egg, Xenopus egg, and the fission yeast Schizosaccharomyces pombe. S. pombe is an excellent system to investigate the changes in the actin cytoskeleton during cell cycle since F-actin patches, F-actin cables and F-actin ring are only visible structures in the cell. The F-actin ring is considered to correspond to the contractile ring in animal cells. It is formed during anaphase in this organism. We have examined process of formation of the F-actin ring by optical sectioning and 3-D reconstitution fluorescence microscopy. In wild type cells, both formation of an aster-like structure composed of F-actin cables and accumulation of F-actin cables was recognized at the medial cortex of the cell during prophase to metaphase. The formation of the aster-like structure seemed to initiate by branching of the longitudinal F-actin cables at a site near the spindle pole bodies which had been duplicated but not yet separated. A single cable extended from the aster and encircled the cell at the equator to form a primary F-actin ring during metaphase. During anaphase, the accumulated F-actin cables were linked to the primary F-actin ring, and then all of these structures seemed to be packed to form the F-actin ring. These observations suggest that formation of the aster-like structure and the accumulation of the F-actin cables at the medial region of the cell during metaphase may be required to initiate the F-actin ring formation. We further examined F-actin structures in both cdc12 and cdc15 early cytokinesis mutants. As a result, Cdc12 seemed to be required for the primary F-actin ring formation during prophase, while Cdc15 may be involved in both packing the F-actin cables to form the F-actin ring and rearrangement of the F-actin after anaphase. In spg1, cdc7 and sid2 septum initiation mutants, the F-actin ring seemed to be formed in order. In large eggs which undergo unilateral cleavage, it has been proposed that localized elevation of cytosolic free calcium ions ([Ca2+]i) at the growing end of the cleavage furrow (CF) triggers CF formation by the activation of myosin ATPase activity through the myosin light chain kinase. However, data regarding the relevance of Ca waves in CF formation is contradictory or incomplete. We have improved the method to visualize Ca wave in dividing Xenopus eggs: we removed the fertilization membrane and labeled the egg surface with rhodamine-wheat germ agglutinin (WGA) to visualize the growing ends of the early CF. We imaged wave type Ca signal with Calcium Green-1 dextran from the animal hemisphere of the egg. In addition to the Ca wave, which is the orchestrated Ca release from a global area of the cell, smaller classes of Ca signal have been described. These are Ca puffs, which are thought to be Ca release from 10-30 of coordinately opened Ca channels, and Ca blips, which are Ca released from single opened Ca channel. These signals were also examined during the furrow formation. Consequently, we could not detect any Ca wave, Ca puff, or Ca blip at the growing end of the CF, although we could detect two Ca waves at later stages of cytokinesis. Furthermore, we lowerd [Ca2+]i in the egg by injection of Ca-chelators. However, it did not affect cleavages of the egg. Therefore, we concluded that Ca signals are not involved in CF formation in Xenopus egg. We also concentrate our study on function of actin-regulatory proteins, including ADF/cofilin family proteins, during cytokinesis using Xenopuseggs and em-bryos. ADF/cofilin family proteins exist in all animals and plants examined and have been shown to be essential. We found that ADF/cofilin family proteins are essential for cytokinesis. Recent studies revealed that ADF/cofilin accelerates turnover of actin filaments both in vitro and in vivo. Most recently, we found a novel actin-regulatory protein which induces disassembly of actin filaments cooperatively with ADF/cofilin. cDNA analysis revealed that this protein is a Xenopus homologue of yeast actin interacting protein 1 (AIP1). Thus, we designated this protein as Xenopus AIP1 (XAIP1). Purified XAIP1 itself binds to pure actin filaments to some extent, but it induces a rapid, drastic disassembly of actin filaments associated with cofilin. Microinjection of this protein into Xenopus embryos arrested development by the resulting actin cytoskeletal disorder. XAIP1 represents the first case of a protein cooperatively disassembling actin filaments with ADF/cofilin family proteins. In addition, we also identified the second protein which functions with ADF/cofilin. That was a Xenopus homologue of cyclase-associating protein (CAP), originally reported as an actin monomer-binding protein. Our biochemical analyses, however, suggest that XCap accelerates nucleotide exchange of actin monomer and induces actin polymerization from actin-cofilin complex at steady state. Publication List:Asano, Y., and Mabuchi, I. (2001). Calyculin-A, an inhibitor for protein phosphatases, induces cortical contraction in unfertilized sea urchin eggs. Cell Motil. Cytoskele.48, 245-261. Arai, R., and Mabuchi, I. (2002). F-actin ring formation and roles of F-actin cables in the fission yeast Schizosaccharomyces pombe. J. Cell Sci. in press. Bernstein, B. W., Painter, W. B., Chen, H., Minamide, L. S., Abe, H. and Bamburg, J. R. (2000). Intracellular pH Modulation of ADF/Cofilin Proteins. Cell Motil. Cytoskel. 47, 319-336. Motegi, F., Arai, R., and Mabuchi, I. (2001). Identification of two type V myosins in fission yeast, one of which functions in polarized cell growth and moves rapidly in the cell. Mol. Biol. Cell 12, 1367-1380. Nakano, K., Sato, K., Morimatsu, A., Ohnuma, M., and Mabuchi, I. (2001). Interactions among a fimbrin, a capping protein, and an actin-depolymerizing factor in organization of the fission yeast actin cytoskeleton Mol. Biol. Cell12, 3515-3526. Nakano, K., Muto, T., and Mabuchi, I. (2001). Characterization of GTP-activating proteins for the Rho-family small GTPases in the fission yeast Schizosaccharomyces pombe. Genes to Cells6, 1031-1042. Noguchi, T., and Mabuchi, I. (2001). Reorganization of actin cytoskeleton at the growing end of the cleavagefurrow of Xenopus egg during cytokinesis. J. Cell Sci.114, 401-412. Noguchi, T., and Mabuchi, I. (2002). Localized calcium signals along the cleavage furrow are not involved in the furrow formation. Mol. Biol. Cell in press. Takahashi, T., Koshimizu, U., Abe, H., Obinata, T. and Nakamura, T. (2001). Functional Involvement of Xenopus LIM-Kinases in Progression of Oocyte Maturation. Dev. Biol.229, 554-567. Toya, M., Motegi, F., Nakano, K., Mabuchi, I., and Yamamoto, M. (2001). Identification and functional analysis of the gene for type I myosin in fission yeast. Genes to Cells6, 187-199. Yamashiro, S., Kameyama, K., Kanzawa, N., Tamiya, T., Mabuchi, I., and Tsuchiya, T. (2001). The gelsolin/fragmin family protein identified in the higher plant Mimosa pudica. J. Biochem. 130, 243-249. Yonemura, I., and Mabuchi, I. (2001). Heterogeneity of mRNAs coding for Caenorhabditis eleganscoronin-like proteins Gene271, 255-259.
Fig. 1 First cleavage of a Xenopus egg. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |