DIVISION OF CELLULAR COMMUNICATION (ADJUNCT)
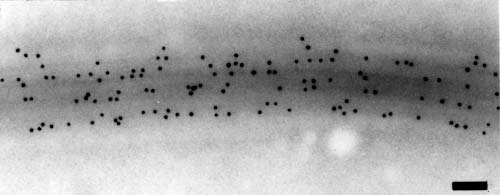
The research in this laboratory is aimed at an understanding of the molecular mechanisms that regulate the assembly and function of cytoskeletal proteins. Specifically, we are currently studying the functional properties of axonemal dynein and actin in Chlamydomonas, an organism ideally suited for genetic and molecular biological studies. I. Function of Multiple Axonemal DyneinsIt is well established that the beating of cilia and flagella is based on sliding movements of outer-doublet microtubules driven by motor proteins dyneins, but how the sliding is converted into axonemal oscillatory bending movement has not been made clear. Recently, various lines of evidence have suggested that dynein is crucially important also in the sliding-bending conversion mechanism. Thus our research effort is now focused on the properties of various dyneins. Biochemical studies by us and other laboratories have established that a single flagellar axoneme contains at least eleven kinds of dynein heavy chains in inner and outer arms. An important question is how different dynein heavy chains differ in function. To answer this question, we have been isolating and characterizing mutants that lack different kinds of axonemal dyneins. During the last ten years, we have isolated as many as 15 genetically different mutants lacking various subsets of dyneins. The isolation of these mutants greatly advanced our understanding of the function and organization of various dyneins within the axoneme, because only three mutants had been known to lack dynein heavy chains before we started mutant isolation. The motility phenotypes of the isolated mutants have indicated that different dynein species differ in function in a fundamental manner. For example, the outer-arm heavy chains are important for flagellar beating at high frequency, whereas the inner-arm heavy chains are important for producing proper waveforms. Indirect evidence also suggests that the force generation properties differ greatly among different heavy chains. Interestingly, the axoneme can beat without some of these heavy chains, but cannot beat if certain combinations of heavy chains are lost. It appears that simultaneous presence of dyneins with different properties is necessary for the axonemal beating. Thus, it should be important to understand the mechano-chemical property of each dynein. To this end, we are currently trying to directly measure the force production in wild-type and mutant axonemes that lack various combinations of dyneins. In addition to exploring the functional properties of individual dyneins, we have been studying the mechanism by which various dyneins are regularly arranged within the axoneme. In particular, we have concentrated our effort on understanding why outer dynein arms are attached to a particular protofilament in outer doublet microtubules, with a constant spacing of 24 nm. Such a regular arrangement of dynein should be important for the axoneme to beat with regular waveforms. Our study has indicated that a protein complex called the outer dynein arm docking complex (ODA-DC) is particularly important. We found that this complex, made up of three different protein subunits, is transported to the axonemes independently of outer arms and attaches on to the microtubule protofilament, providing the docking site for outer arms. Interestingly, the periodicity of 24 nm appears to originate from the periodicity in ODA-DC binding (Fig. 1). An attractive hypothesis is that ODA-DC has a tendency to longitudinally associate with each other and serves as a molecular ruler with a 24 nm spacing. Our study has also shown that the outer doublet microtubule contains an unidentified structure that can specify the ODA-DC binding site among different protofilaments of a single outer doublet. We are currently trying to elucidate this structure, as well as to prove the molecular ruler hypothesis. II. Function of Actin and an Actin-related Protein in ChlamydomonasThe inner dynein arms are known to contain actin as a subunit. Hence the two independent motility systems of eukaryotes - the actin-based and microtubule-based motility systems - should somehow cooperate in the inner arm dynein although the function of actin in dynein arms is totally unknown at present. We have found that the mutant ida5, lacking four out of the seven subspecies of inner-arm dyneins, has a mutation in the actin-encoding gene. Intriguingly, Chlamydomonas has been known to have only a single gene of conventional actin, and the mutant ida5 was found to express no conventional actin at all. On close inspection, the cytoplasm and axonemes of this mutant were found to contain a novel actin-like protein (NAP) which displays exceptionally low homology (64%) to conventional actin. The mutant ida5is deficient in the formation of the fertilization tubule and thus has a low mating efficiency. However, it displays normal cell division and grows as rapidly as wild type, possibly because NAP can substitute for actin in important cellular functions. Thus conventional actin and NAP may overlap in some, but not all, cellular functions. It is interesting to note that NAP is expressed in significant amount only in the mutant lacking actin; i.e., the expression of NAP appears to depend on the presence of actin. We are currently investigating how such regulation takes place. What is the function of NAP in wild-type cells? Recently, we found NAP is expressed even in wild type, specifically during the re-flagellation process after flagellar amputation. The expression of NAP mRNA precedes that of actin mRNA. Interestingly, cell's ability to re-grow flagella is greatly impaired when ida5 is transformed with a chimeric actin gene consisting of a 5'-UTR sequence derived from the NAP gene and a sequence that codes for conventional actin. NAP and actin mRNA are expressed simultaneously in this mutant, suggesting that the prior expression of NAP is important for efficient flagellation. Phylogenetic analysis revealed that NAP homologs exist in at least three species in Volvocales, forming a distinct gene family of divergent actin. Taken together, NAP may play a specific role in the flagellation mechanism in various species of Volvocales. Publication List:Hayashi, M., Hirono, M., and Kamiya, R. (2001) Recovery of flagellar dynein function in Chlamydomonas actin/dynein-deficient mutant upon introduction of muscle actin by electroporation. Cell Motil. Cytoskeleton49, 146-153. Inaba, K. (2001) High level of protein ubiquitination in ascidian sperm. In The biology of ascidians. (Sawada, Yokosawa, Lambert, eds,), pp.74-79.,Springer-Verlag Itoh, A., Inaba, K., Fujinoki, M., and Morisawa, M. (2001) Motility-associated and cyclic AMP-dependent protein phosphorylation in the sperm of the chum salmon, Oncorhynchus keta. Biomed. Res.22, in press. Kho, K. H., Tanimoto, S., Inaba, K., Oka, Y., and Morisawa, M. (2001) Transmembrane cell signaling for the initiation of trout sperm motility: Roles of ion channels and membrane hyperpolarization for cyclic AMP synthesis. Zool. Sci.18, 919-928. Padma, P, Hozumi, A, Ogawa, K., and Inaba, K. (2001) Molecular cloning and characterization of a thioredoxin/NDPK related dynein intermediate chain from the ascidian, Ciona intestinalis. Gene275, 177-183. Takada, S., Wilkerson, C. G., Wakabayashi, K., Kamiya, R., and Witman, G. B. (2001) The outer dynein arm docking complex: composition and characterization of a subunit (Oda1) necessary for outer arm assembly. Mol. Biol. Cell in press. Wakabayashi, K., Takada, S. Witman, G. B., and Kamiya, R. (2001) Transport and arrangement of the outer-dynein-arm docking complex in the flagella of Chlamydomonasmutants that lack outer dynein arms. Cell Motil. Cytoskeleton48, 277-286. Yanagisawa, H. and Kamiya, R. (2001) Association between actin and light chains in Chlamydomonas flagellar inner-arm dyneins. Biochem. Biophys. Res. Commun.288, 443-447. Yoshimura, K. and Kamiya, R. (2001) The sensitivity of Chlamydomonas photoreceptor is optimized for the frequency of bodily rotation. Plant Cell Physiol.42, 665-672.
|
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |