DIVISION OF BIOLOGICAL REGULATION AND PHOTOBIOLOGY
(ADJUNCT)
Professor (adjunct):
Kimiyuki Satoh
Associate Professor (adjunct):
Hirokazu Kobayashi
Research Associates:
Satoru Tokutomi
Noritoshi Inagaki
Graduate Students:
Kyoichi Isono *
Yoshihiro Narusaka *
Tatsuya Tomo *
Akira Matsumoto *
(* from Okayama University)
From an energetic point of view, the conversion of light energy into chemical energy in photosynthesis is the most important biological process on earth. The highly efficient energy conversion in this process is ensured by the highly ordered organization of molecules in the photochemical reaction center, in a physical, chemical and biological sense. The project in this division is aiming to elucidate the organization of photosystem II reaction center of oxygenic photosynthesis which has a unique property to generate a strong oxidant for utilizing the water molecule as electron donor. In the first approach, molecular organization of the photosystem II reaction center, which has been identified in our study, will be analyzed by several methods which include crystallographic analysis, chemical modification & cross-linking analysis and optical & EPR spectroscopies. Structure-functional analysis will also be conducted for the reaction center using random and site-directed mutagenesis for transformable algae, Synechocystis PCC 6803 and Chlamydomonas reinhardtii. The principal target of these analyses will be the structure and molecular environment of P-680, the primary donor, which determine the redox potential of this unique photosystem.
In the second approach, the effort will be focused on the elucidation of molecular mechanism of light-regulated metabolic turnover of a subunit of the photosystem II reaction center, D1 protein. Some of unique steps are involved in this process; i. e., photo-damage of the function of photochemical reaction center, specific degradation of the impaired protein subunit, Iight-regulated gene expression at the translational level, post-translational cleavage of the C-terminal extension and the incorporation of cofactors and subunits into multi-component pigment-protein complexes.
I. Structural organization of photosystem II reaction center
The structure and molecular interactions of the primary donor in the photo-system II reaction center have been investigated by detecting light-induced FT-IR difference spectra upon the formation of its triplet state. From the band positions of the keto and carbomethoxy C=0 stretches, the hydrogen-bonding properties of the two chlorophylls of P-680 were found to be asymmetrical, in one chlorophyll (P1) both the keto and carbomethoxy C=0 groups form hydrogen bonds, while in the other chlorophyll (P2) the keto C=0 is not hydrogen-bonded whereas the carbomethoxy C=O probably is hydrogen-bonded. Considering the orientation of P-680 analyzed by EPR and the structure of bacterial reaction center determined by X-ray crystallography, together with the sequence homology between the D1 and D2 subunits of photosystem II and the L and M subunits of purple bacteria, a model of the structure of P-680 and its interactions with apoproteins has been proposed (Noguchi, T., Inoue, Y. and Satoh, K., 1993, Biochemistry,32, 7186-7195). Site-directed modification using Chlamydomonas reinhardtii of the amino acid side chains on D1 protein presumably involved in the hydrogen-bonding interaction (Ser-191 and Thr-192) has been succeeded and the analysis of these mutants is now in progress in order to prove this hypothesis.
Chemical cross-linking analysis has also been conducted for the isolated photosystem II reaction center to analyze the gross structure; i. e., the nearest neighbors of the D1 and D2 subunits and amino acid residues in cross-contact in the reaction center complex.
II. Dynamic aspects of the organization of photosystem II reaction center
(1) The D1 subunit of photosystem II reaction center has a C-terminal extension consisting of 8-16 amino acids. Post-translational removal of this part of the protein is essential for the manifestation of oxygen-evolving function in photosystem II. The enzyme involved in the processing has been identified and the partial amino acid sequences have been determined for spinach. Based on these data, a gene coding for the enzyme has been identified and sequenced (paper in pre paration).
(2) The recognition signal on substrate was analyzed for the C-terminal processing protease using substituted synthetic oligopeptides corresponding to the C-terminal sequence of D1 precursor protein. The efficiency of these synthetic oligopeptides, both as substrate and as inhibitor for the C-terminal cleavage, were examined. The result suggests that the presence of a hydrophobic amino acid side chain at the -2 position, in addition to the secondary structure around the cleavage site, is crucial in the recognition (Taguchi, F., Yamamoto, Y. and Satoh, K., J. Biol. Chem., in press).
(3) The synthesis of D1 precursor protein is regulated by light at the stage of translation. The mechanism of this light regulation was analyzed using isolated pea chloroplasts. The isolated chloroplasts was shown to accumulate translation intermediate(s) of D1 protein in the presence of externally added ATP. The result suggested that the translation of D1 protein at the specific stage of polypeptide elongation requires factor(s) caused by illumination, in addition to energy supply.
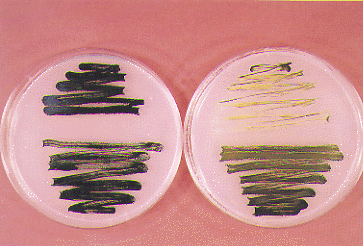
(4) Photo-tolerant mutants of an unicellular cyanobacterium Synechocystis PCC 6803 were obtained by in vitro random mutagenesis of psbAII (gene for D1 protein) by PCR under a condition for reduced fidelity of amplification, in order to analyze the damage-repair cycle of D1 protein in photoinhibition of photosystem II reaction center. The mechanistic analysis of photo-tolerance caused by these amino acid displacements on D1 protein is now in progress. Figure shows a photo-tolerant mutant (lower), together with the control strain (upper), grown at the photon flux density of 50µE (left) and those after exposure to 320µE (right).
(5) Synthesis of some photosystem proteins are regulated by light signals received by phytochrome. Molecular mechanism of the photoreception of phytochrome were studied by circular dichroism and resonance Raman spectroscopies. The former shows that the phototransformation of pea phytochrome from Pr to Pfr induces unfolding of a -helix in the 6-kDa N-terminal domain which is requisite for its function. The latter proposes that a proton associated to the chromophore in Pr migrates to the apoprotein during a phototransformation process(s) to the bleached intermediate, which presumably triggers conformational changes in the molecule.
Publication List:
Deforce, L., Tokutomi, S. and Song, P.-S. (1994) Phototransformation of pea phyiochrome A induces an increase in a -helical folding of the apoprotein: Comparison with a monocot phyiochrome A and CD analysis by different methods. Biochemistry33, 4918-4922.
Mizutani, Y., Tokutomi, S. and Kitagawa, T. (1994) Resonance Raman spectra of the intermediates in phototransformation of large phytochrome: Deprotonation of the chromophore in the bleached intermediate. Biochemistry33, 153-158.
Nozawa, T., Kobayashi, M., Wang, Z.-Y., Itoh, S., Iwaki, M., Mimuro, M. and Satoh, K. (1994) Magnetic circular dichroism investigation on chromophores in reaction centers of photosystem I and II of green plant photosynthesis. Spectrochimica Acta,. in press.
Tomo, T. and Satoh, K. (1994) Nearest neighbor analysis of D1 and D2 subunits in the photosystem II reaction center using bifunctional cross-linker, hexamethylene diisocyanate. FEBS Lett.351, 27-30.
National Insitute for Basic Biology