《 基礎生物学研究所要覧 》形質統御実験施設
|
| DIVISION OF SPECIATION MECHANISMS I |
|
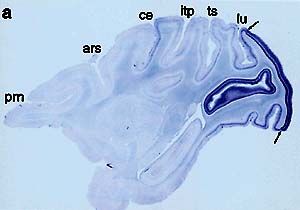
1.大脳皮質の領野特異性とその進化(1)大脳皮質領野の決定機構大脳皮質領野が発生的にどのようにして決定されるのかということについては,従来より2つの異なる考え方がある。一つは,将来大脳皮質を将来構成する細胞が腦室の分裂層にある時にすでにその運命が決定されているという考えと,今一つは,視床からの入力によって視覚野,聴覚野等への領域特異性が決定されるという考え方である。この10年余りの間に,げっ歯類を材料に用いた研究に於いては,レトロウイルスベクターを用いた実験や移植実験から得られていた後者を示唆する考え方から,大脳皮質の領域(この場合領野よりは広い)に特異的に発現する遺伝子が幾つか調べられ,それが視床の入力とは独立にその発現が制御されていることが示されるという考え方の大きな転換があった。しかし,大脳皮質領野の決定がどの程度まで遺伝的にプログラムされており,どの程度まで環境入力によって可変的かは,未だ結論を見ていない。 (2)大脳皮質の進化大脳皮質は,哺乳類,殊にヒトで最も顕著に発達している。例えば,神経細胞を作る分裂組織や海馬等では,体重比で補正して,原始的哺乳類である食虫類とヒトでは4〜5倍程度の差しか無いにも拘わらず,大脳皮質では200倍もの差がある。このことは,哺乳類の脳機能の進化に於いて,大脳皮質の進化が極めて重要であることを示している。ネズミと霊長類の比較解剖学的な対象は,大脳皮質以外の脳構造については,95%近くの対応がついているが,大脳皮質については,逆に殆ど対応がついていない状況である。最近のヒト遺伝子のドラフト配列の発表によっても,ヒトとマウスでは,遺伝子数は殆ど変化していないとされている。にも拘わらず,どのようにしてこうした大脳皮質領野の急速な拡大がもたらせられたのか,非常に興味深い。 (3)霊長類の大脳皮質領野特異的に発現する遺伝子私達は,上述した大脳皮質領野特異性の発生と進化の未解決の問題を分子細胞レベルから解明する為には,大脳皮質の発達した霊長類の大脳皮質領野に特異的に発現する遺伝子を分離し解析することが極めて有効と考え,研究を開始した。 先ず,マクロアレイ法により,1088遺伝子中,ヒトの3領野(前頭葉,運動野,後頭葉)に於いて,どの程度の遺伝子発現の差異が見られるのか検討した(那波新潟大脳研教授との共同研究)。その結果,個体差を平均化した上で領野間の差を比較すると,最大3〜4倍の差異を示すものが1つ,2〜3倍のものが1つある以外は,全て2倍以内の差異しかなかった。従って,大脳皮質の遺伝子発現は,意外な程領野間での差がないことが分った。 しかし,この結果は,領野間での発現パターンが異なるものが存在しないということを意味するものではない。数は少なくとも領野間で顕著な発現の差を示す遺伝子が存在する可能性はある。そこで,Differential Display法を用いて,霊長類(マカカ属)の大脳皮質の代表的的領野間(前頭葉,運動野,側頭葉,視覚野等)で発現に顕著な差が見られる遺伝子を探索した。その結果,領野間で最大10倍以上の差のある3個の遺伝子を見い出した。そのうちの一つは,視覚野に特異的に発現する遺伝子occ1(occipital1)であり,他の一つは,運動野特異的に発現する遺伝子(gdf7)である。第3番目の遺伝子については,現在解析中である。 例えば,occ1は,一次視覚野に顕著に発現がみられ,2次視覚野では,急激にその発現が低下し,更に前部に移行するに従って,その発現量は急速に低下する(図1)。これは,前述したブロードマンの領野に厳密に対応する発現パターンを示すおそらく最初の例である。従って,occ1は,大脳皮質の視覚野がどのように発生と進化的制御を受けているのかを明らかにする上で,極めて有効なマーカーとなり得ると考え解析を進める予定である。更に興味深いことに,この遺伝子は,片眼にテトロドトキシン(TTX)を注入して,網膜の電気的活動を遮蔽すると,視覚野の眼優位性カラムのうち,TTXを注入した眼より入力を受けるカラムに於いてのみ,顕著な低下を見せる。 私達は,occ1 やgdf7等の大脳皮質の領域特異的な顕著な発現パターンを示す遺伝子は,約3万遺伝子の内でも,おそらく30個以上は無いと推測しているが,現在,RLCS法により20個程の遺伝子を分離しており,このような遺伝子の網羅的解析から,哺乳類の大脳皮質の発生と進化の様式を明らかにしたいと考えている。
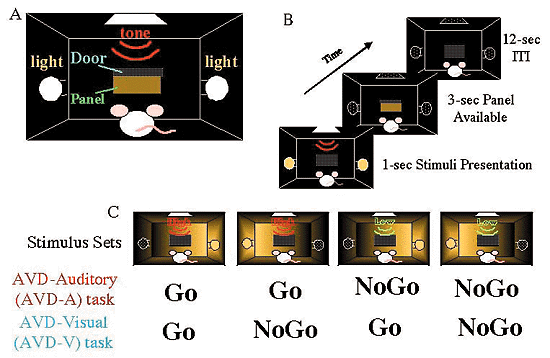
2.学習行動下での遺伝子発現大脳皮質の機能を解析するには,電気生理的方法やイメージング等種々の方法が考案されているが,各々に時間分解能,空間分解能の長所と短所がある。私達の研究室では,c-Fos等の遺伝子発現を指標に,特に細胞レベルでの脳神経回路の結合様式の変化を研究している。用いている学習システムは2つである。一つは,京都大学文学部の櫻井芳雄教授との共同研究として行っている視聴覚弁別学習課題である(図2)。高音と低音,左右の光源の何れか一つを学習の刺激条件として,他を対照刺激としてランダムに呈示し,餌報酬により訓練したラットに於いて,例えば音刺激条件下でのc-Fosの聴覚野と視覚野に於ける発現量を比較したところ,聴覚野で有意にc-Fos発現の増大が見られた。更に,この課題依存的なc-Fosの発現が興奮性の神経細胞にのみ見られることを明らかにし,電気生理学方法や従来のイメージング法では難しい細胞レベルでの神経回路網の変化を知ることが可能であることを示した。 今一つは,私達の研究室で開発した,ホイール走行システムである。これは,ホイール上の足場の形を変化させて回転したときマウスがその形に応じて走行できる様になるのに必要な脳内に於ける神経回路を調べるものであり,手続き記憶の脳内経路を細胞レベルで明らかにすることを目指しているが、線状体の介在神経のサブクラスによって、パターン変化時のc-Fos発現が異なるという興ある現象を見い出している。この知見も従来の電気生理学やイメージングでは、知られていないものであり、行動解析と結び付けた遺伝子発現の手法が有効であることを示している。
参考文献
|
| Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) | ||