
|
|
DIVISION OF THEORETICAL BIOLOGY |
| Associate Professor: | MOCHIZUKI, Atsushi |
| NIBB Research Fellow: | TOHYA, Shusaku |
| Postdoctoral Fellow: | FUJITA, Hironori |
| Graduate Students: | AYABE, Yoshiko |
| YAMAGAMI, Ayumi | |
| IMAMURA, Hisako | |
| Secretary: | UMEBAYASHI, Hiromi |
I. Mathematical approach to biological phenomenaWe are studying biological phenomena by using mathematical models. The method gives us an integrative understanding for behavior of complex systems in biology including gene regulatory networks. The mathematical models are also useful to understand pattern formation in development. Study of the mechanisms responsible for morphological difference between species is an important research focus of the current developmental biology. Theoretical studies would be useful in identifying candidates of cell-cell interaction that are likely to be responsible for the systems in real organisms. II. A theoretical study for gene regulatory networks and analysis of differentiated cell-statesThe highly complex behavior of organisms is based on the interactive regulation of genes, functioning together to create a gene network. The regulation of gene expression levels is dependent on the concentrations of transcriptional regulators.Typically, multiple transcription factors can bind to the regulatory region of a gene; each factor may independently affect the gene expression levels. In this manner, the regulatory region of gene serves as an integration point, performing a logical computation of these transcription factors to determine the resulting gene expression levels. In addition, once translated, the encoded protein may control the transcription of other genes. The complexity of gene networks has been considered to be a problem that may be solved from a theoretical point of view. Since the proposal of the Boolean model by Kauffman, multiple mathematical models have been invented and analyzed to study complex gene networks. There are two general classes of network models, discrete and continuous models.Continuous models are likely more realistic than discrete models as a representation of biological systems. However, the dynamical behavior of continuous models is analyzed much less than that of discrete models. In this paper, I have developed a model for gene networks in a framework of continuous time and state variables. I analyze steady states appearing in this system in general way. The steady states specify distinct cell states defined by patterns of gene activity. I introduce three types of steady states observed in a continuous time and space model, whose values and stability can be evaluated from the structure of gene regulatory interactions. The expected numbers of steady states in a randomly generated gene network were derived in exact forms. I determined the distribution of the steady states in a randomly generated network. The results were surprising; the number of steady states in a random gene network does not depend on gene number. In addition, the number of steady states is very small; the number of RSPs, in which the activity of each gene is either on or off, is only one. Most randomly generated gene networks will have only a few (or no) steady states, despite the presence of many genes in the system. Thus, increases in the number of genes do not augment the number of steady states for gene activity. The gene expression steady states may correspond to differentiated cell type identities. Thus, this result implicates that the number of genes is not the driving force increasing the diversity of differentiated cell types or the complexity of organisms. |
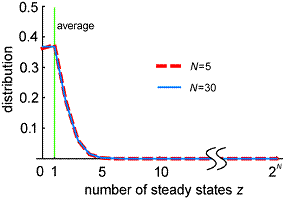 |
Figure 1. Distribution of the number of regular stationary points (RSPs). The distribution demonstrates a case that the regulation of each gene in each domain is independent. The two lines show the results of gene number N is 5 and 30. The distribution does not hardly depend on gene number |
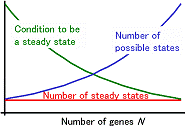 |
Figure 2. Intuitive explanation for the independence of steady state number on gene number. The number of domains 2N, the maximum number of RSPs, increases exponentially with gene number N. The probability for each domain to contain an RSP, however, decreases with gene number. The two effects are opposing and equivalent. Thus, the effect of N on the number of steady states disappears. |
|
In reality, multi-cellular organisms have a wide variety of differentiated cells. Why, then, do the networks in actual organisms have multiple steady states? It seems likely that the networks within actual organisms are quite biased from the average randomly generated network. A gene network including a large number of self-regulatory genes has a large number of steady states. Thus, self-regulation within a gene network may be the main source of cell type diversity in actual organisms. III. Mathematical models for pattern formation of leaf vascular networks.The vascular system of plant is a network of bundles that connects within major organs. The development or differentiation of vascular system is one of the most important subjects in botany, and a lot of studies have focused the problem. In dicot systems, veins in a leaf are usually diverse in their size and they can be classified distinctively by their appearance. The largest vein located at the center of a leaf is called midvein (or primary vein), which are generated first. Veins differentiate progressively in an early developmental stage of leaf. A vein, which is continuous with stem vascular bundle, extends starting from proximal edge to distal end of the leaf, and then become a midvein. Secondary veins are generated from branches from the midvein and are thinner than the midvein. By repeating branching and extension, higher order veins are generated sequentially with growth of leaf and the reticulate network system is formed. Although a lot of experimental studies have attempted to identify the mechanism of vascular patterning in leaves, it is still a problem to solve. Jacobs revealed the importance of auxin for vascular differentiation. Auxin is one of the major plant hormones. In the higher plant, indole acetic acid (IAA) is one of the most important auxin chemicals. Auxin is diffusible molecule; however, it is considered that auxin is also transported along the particular direction, usually from apical to basal in a stem, from distal to proximal in a leaf. It is called auxin polar transport. In a leaf, auxin is considered to be produced in apical margins of leaves and transported toward the proximal regions. Many researchers considered that such auxin flow play important role in vascularization of plant. Three hypotheses have been proposed so far to explain the leaf venation pattern formation. One is auxin canalization hypothesis proposed by T. Sachs that is based on the assumption of the positive feedback regulation between auxin flow and flow capacity. Auxin is a diffusible plant hormone of small molecule and has important roles in various developmental events including vascular differentiation in plants. In this hypothesis, auxin pathways are expected to be generated from a uniform field and are extended in a sequential manner. The second is activator-inhibitor type reaction-diffusion hypothesis proposed by H. Meinhardt that is based on two interactive factors of weakly diffusible activator and strongly diffusible inhibitor. We proposed the third hypothesis, substrate-depletion type reaction-diffusion model, that is based on two interactive factors of strongly diffusible auxin resource and its consumer. |

|
Figure 3. Branching pattern generated by computer simulation based on substrate-depletion type reaction-diffusion model. Veins grow toward the margin with repeating branching. In the final state, regular interval branching patterns are formed. |
|
On the other hand, we tried to integrate the canalization hypothesis and the experimental knowledge. we introduce auxin transport by PIN1 efflux carrier into the auxin canalization hypothesis. So we investigated a model based on the assumption of the positive feedback regulation between auxin flow and PIN1 localization. The substrate-depletion type reaction-diffusion model can regenerate growth and branching patterns. On the other hand, the canalization model can generate closed circuits of leaf veins under a condition. Both two mechanisms seem to be responsible for the leaf vein formation. |
 |
Figure 4. Leaf vein pattern generated by computer simulation based on canalization model. Veins grow toward the margin with repeating branching. The final patterns are similar to ones observed in maple leaves. |
Publication ListMochizuki, A. (2005) An Analytical Study of The Number of Steady States in Gene Regulatory Networks. J. Theor. Biol. (In Press). Tohya, S. & Mochizuki, A. (2005) A new model for leaf venation pattern. J. Theor. Biol. (In Press). Fujita, H. & Mochizuki, A. (2005) Pattern formation by the positive feedback regulation between flow of diffusible signal molecule and localization of its carrier. J. Theor. Biol. (In Press). Tohya, S., Mochizuki, A. and Iwasa, Y. (2003) Random cell sorting can form cone mosaic patterns in fish retina and explain the difference between zebrafish and medaka. J. Theor. Biol.221, 289-300. Shoji, H., Mochizuki, A., Iwasa, Y., Hirata, M., Watanabe, T., Hioki, S. & Kondo, S. (2003) Origine of directionality in the fish stripe pattern. Dev. Dyn. 226, 627-633. Ryohji, T., Mochizuki, A. & Iwasa, Y. (2003) Possibility of Tissue Separation Caused by Cell Adhesion. J. Theor. Biol.221, 459-474. |
 |