DIVISION OF PLANT DEVELOPMENTAL GENETICS
Diversity of plant form is mostly attributable to variation of leaf and floral organs, which are modified leaves. The leaf is the fundamental unit of the shoot system, which is composed with leaf and stem. So the leaf is the key organ for a full understanding of plant morphogenesis. However, the genetic control of development of these shapes had remained unclear. Recently, studies of leaf morphogenesis has been in a turning point, after our successful application of the techniques of developmental and molecular genetics to it, using model plants, Arabidopsis thaliana (L.) Heynh (reviewed in Tsukaya 2002a, b). In a determinate organ, a leaf, number of leaf cells is not necessarily reflected on leaf shape or, in particular, leaf size. Genetic analyses of leaf development in Arabidopsis shows that a compensatory system(s) act in leaf morphogenesis and an increase of cell volume might be triggered by a decrease in cell number. Thus, leaf size is, at least to some extent, uncoupled from the size and number of cells by the compensatory system(s). Based on these facts, a new perspective on understanding of mechanisms for leaf morphogenesis, Neo-Cell theory, is proposed (Tsukaya, 2002a; Tsukaya, 2003). Neo-Cell theory is the Cell theory which stipulated positional cooperation of the behavior of cells. In short, Neo-Cell theory hypothesizes as follows: cells are the unit of morphogenesis; however, each cell is also controlled by factors that govern the morphogenesis of the organ of which the cells are a part. Focusing on mechanisms that govern polarized growth of leaves in a model plant, Arabidopsis thaliana, we found that the two genes act independently to each other on the processes of polar growth of leaves: the ANGUSTIFOLIA (AN) gene regulates width of leaves and the ROTUNDIFOLIA3 (ROT3) gene regulates length of leaves. The AN gene controls the width of leaf blades and the ROT3 gene controls length. Cloning of the AN gene (Kim et al., 2002) revealed that the gene is a member of CtBP (C-terminal binding protein) gene family which had been found from animal kingdom. In the animal kingdom, CtBPs self-associate and act as a co-repressors of transcription. We found that also the AN protein can self-associate in yeast two-hybrid system (Kim et al. 2002). Furthermore, microarray analysis suggested that the AN gene might regulate the expression of certain genes, for example, the gene involved in formation of cell walls, MERI5. We also found that the abnormal arrangement of cortical MTs in an-1 mutant leaf cells appeared to account entirely for the abnormal shape of the cells. It suggested that the AN gene might regulate the polarity of cell growth by controlling the arrangement of cortical MTs. The ROT3 gene was cloned by us in 1998. Transgenic experiments proved that the ROT3 gene regulates leaf-length without affect on leaf-width via biosynthesis of steroids (Kim et al., 1999). In relation to it, we recently revealed that a steroid hormone, brassinosteroid, controls both proliferation and expansion of leaf cells (Nalaya et al., 2002). In addition, by studying role of light condition in leaf development, we found that the length of leaf petioles was related exclusively to genetic control of the length of individual cells in all leaf morphological mutants examined. In contrast, both the size and the number of cells were affected by the mutations examined in leaf blades. Studies of the roles of light signal perception in leaf development are now underway. Apart from the aspects of leaf expansion, we also analyzed genes for identification of leaf primordia. The AS1, AS2 and BOP genes are needed for determinate growth of leaf. Molecular and anatomical analysis of the AS2 gene has been carried out in collaboration with a research team of Prof. Machida, Nagoya University (Endang et al., 2001; Iwakawa et al. 2002). AS2 gene is a member of a novel gene family, including 42 putative genes in Arabidopsis genome, encoded proteins with cysteine repeats and a leucine zipper (Iwakawa et al. 2002). The gene product also represses the expression of class 1 knox homeobox genes in leaves (Endang et al. 2001; Iwakawa et al. 2002), as known for AS1. We also isolated and analyzed a novel mutant, bop (blade-on-petiole) mutant, which strongly enhances the as2 phenotype, in collaboration with a reaseach team of Prof. Nam, POSTECH, Korea (Ha et al. 2003). Three class I knox genes, namely, KNAT1, KNAT2 and KNAT6, were misexpressed in the leaves of the bop1-1 mutant as in those of the as2 mutant. The bop1 single mutant results in ectopic, lobed blades along the adaxial side of petioles of the cotyledon and foliage leaves. We, thus, suggest that BOP1 promotes or maintains a developmentally determinate state of leaf cells through regulation of class I knox1 genes (Ha et al. 2003). On the other hand, we are trying to identify molecular mechanisms which distinguish developmental pathway of leaves from that of shoots by studying tropical plants with 'fuzzy' morphology. For such purposes, we introduced tropical plants having queer developmental program for leaf morphogenesis, namely, Chisocheton, Guarea and Monophyllaea, as materials for the studies. The indeterminate compound leaves of members of the genus Chisocheton in Southeast Asia and of the genus Guarea in the New World and Africa, in the family Meliaceae, are unique and can develop indeterminately as a result of the activity of the leaf apical meristem, which can function very similarly to a shoot apical meristem. We performed a molecular phylogenetic study of these genera, and the result suggested that indeterminate program in the leaves of members of these two genera might have evolved only once in Meliaceae (Fukuda et al. 2003). In addition, we are interested in environmental adaptation of leaves, from view point of biodiversity. For example, thermograph analysis of function of downy bracts of a so-called snowball plant, Saussurea medusa that habits in alpine zone of Yunnan, China, revealed that the downy bracts of S. medusa have two functions: thermal insulation to protect the inside of flowers and the accumulation of heat on the upper surfaces of the inflorescence (Tsukaya et al. 2002a). Anatomical analyses of various types of morphological adaptation of leaves against certain kinds of environments were also performed (Tsukaya, 2002d, e, f, g). In particular, leaf index, relative length of leaf to width, is also the most diverse factor of leaf shape, and is affected by environmental factors in some plants (Tsukaya, 2002c; Tsukaya et al. b). Are AN and ROT3 genes are involved in regulation of adaptive change of leaf index in natural condition? Are these genes the responsible for evolution of rheophytes? So called "Evo/Devo" study of leaf morphogenesis is also one of our research project in NIBB. Publication List:Fukuda, T., Yokoyama, J. and Tsukaya, H. (2003) The evolutionary origin of indeterminate leaves in Meliaceae: phylogenetic relationships among species in the genera Chisocheton and Guarea, as inferred from sequences of chloroplast DNA. Int. J. Plant Sci. 164, 13-24. Ha, C.-H., Kim, G.-T., Kim, B.-C., Jun, J.-H., Soh, M.-S., Ueno, Y., Machida, Y., Tsukaya, H. and Nam, H.-G. (2003) The BLADE-ON-PETIOLE gene controls leaf pattern formation through regulation of meristematic activity. Development 130, 161-172. Iwakawa, H., Ueno, Y., Semiarti, E., Onouchi, H., Kojima, S., Tsukaya, H., Hasebe,M., Soma, T., Ikezaki, M., Machida,C., and Machida, Y. (2002) The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana, required for formation of a symmetric flat leaf lamina, encodes a member of a novel family of proteins characterized by cysteine repeats and a leucine zipper. Plant Cell Physiol. 43, 467-478. Kim, G.-T., Shoda,K., Tsuge, T., Cho, K.-H., Uchimiya, H., Yokoyama, R., Nishitani, K. and Tsukaya, H. (2002) The ANGUSTIFOLIA gene of Arabidopsis, a plant CtBP gene, regulates leaf-cell expansion, the arrangement of cortical microtubules in leaf cells and expression of a gene involved in cell-wall formation. EMBO J. 21, 1267-1279. Kim, G.-T. and Tsukaya, H. (2002) Regulation of the Biosynthesis of Plant Hormones by Cytochrome P450s. J. Plant Res. 115, 169-177. Nakaya, M., Tsukaya, H., Murakami, N., and Kato, M. (2002) Brassinosteroids control the proliferation of the leaf cells in Arabidopsis thaliana. Plant Cell Physiol. 43, 239-244. Stoynova-Bakalova, E., Petrov, P., and Tsukaya, H. (2002) Cell division and cell enlargement in isolated Cucurbita cotyledons grown in darkness and in light. J. Plant Res. 115, 375-380. Tsukaya, H. (2002a) Interpretation of mutants in leaf morphology: genetic evidence for a compensatory system in leaf morphogenesis that provides a new link between Cell and Organismal theory. Int. Rev. Cytol. 217, 1-39. Tsukaya, H. (2002b) Leaf Development, The Arabidopsis Book, eds. C.R. Somerville and E.M. Meyerowitz, American Society of Plant Biologists, Rockville, MD, doi/10.1199/tab.0072, http://www.aspb.org/downloads/arabidopsis/tsukaya.pdf Tsukaya, H. (2002c) The Leaf Index: Heteroblasty, Natural Variation, and the Genetic Control of Polar Processes of Leaf Expansion. Plant Cell Physiol. 43, 372-378. Tsukaya, H. (2002d) Morphological adaptation of bracts in Sino-Himalayan plants that bloom at low temperatures - Downy plants and glasshouse plants - Newslet. Him. Bot. No. 30, 1-7. Tsukaya, H. (2002e) Molecular mechanisms related to the evolution of leaf shape, with a special emphasis on leaf index. In Himalayan Botany in the Twentieth and Twenty-first Centuries. Edited by Noshiro, S. and Rajbhandari, K.R. pp. 152-154, The Society of Himalayan Botany Tokyo, Tokyo. Tsukaya, H. (2002f) Optical and anatomical characteristics of bracts from the Chinese “Glasshouse” plant, Rheum alexandrae Batalin (Polygonaceae), in Yunnan, China. J. Plant Res. 115, 59-64. Tsukaya, H. (2002g) Leaf anatomy of a rheophyte, Dendranthema yoshinaganthum (Asteraceae), and of hybrids between D. yoshinaganthum and a closely related non-rheophyte species, D. indicum. J. Plant Res.115, 329-333. Tsukaya, H. (2003) Organ shape and size: a lesson from studies of leaf morphogenesis. Curr. Opin. Plant Biol. 61, 57-62. Tsukaya,H., Fujikawa, K. and Wu, S.-G. (2002a) Thermal Insulation and Accumulation of Heat in the Downy Inflorescences of Saussurea medusa (Asteraceae) at High Elevation in Yunnan, China. J. Plant Res. 115, 263-268. Tsukaya, H., Kozuka, T. and Kim, G.-T. (2002b) Genetic control of petiole length in Arabidopsis thaliana. Plant Cell Physiol. 43, 1221-1228. 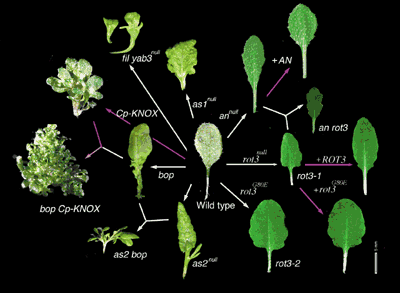
Leaf shape control by various genes in Arabidopsis thaliana. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |