DIVISION OF BIO-ENVIRONMENTAL SCIENCE
1) Graduate School of Yokohama City University Synthetic chemicals found in the environment have the capacity to disrupt endocrine system development and function in both wildlife and humans. This has drawn public concern since many of these chemicals may bind to estrogen receptors (ER) and evoke estrogenic effects. Early evidence that estrogenic chemicals could pose a threat to human health during development came from studies of diethylstilbestrol (DES), which was used to prevent premature birth and spontaneous abortion. Laboratory experiments have demonstrated that exposure of animals to sex hormones during perinatal life can cause permanent and irreversible alterations of the endocrine and reproductive systems as well as the immune system, nervous system, bone, muscle, and liver in both sexes. Although many of these chemicals may bind to ER and evoke estrogenic effects in wildlife and humans, the effects of estrogen are not well understood even now. Thus, understanding the effects of sex hormones at the molecular level, especially during development, is very important to resolve these problems.
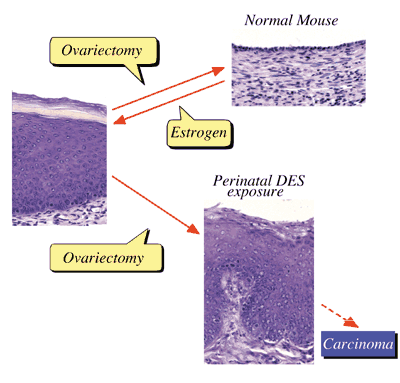
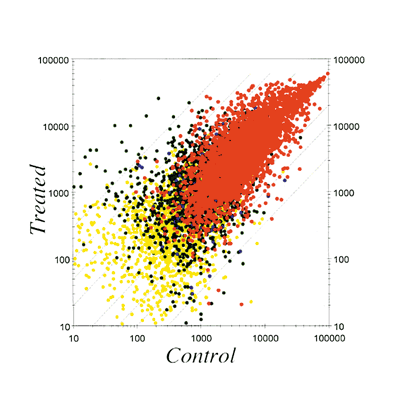
I. Estrogen-induced irreversible changesPerinatal sex-hormone exposure has been found to induce lesions in reproductive tracts in female mice. The possible relevance of the mouse findings to the development of cancer in humans has been emphasized. In the early seventies, a close correlation between occurrence of vaginal clear cell adenocarcinoma in young women and early intrauterine exposure to DES was demonstrated. Many chemicals released into the environment have the potential to disrupt endocrine function in wildlife and humans. Some of these chemicals induce estrogenic activity by binding to ER. The neonatal mouse model has been utilized especially to demonstrate the long-term effects of early sex hormone exposure on the female reproductive tract. Neonatal treatment of female mice with estrogens induces various abnormalities of the reproductive tract: ovary-independent cervicovaginal keratinization, adenosis, uterine hypoplasia, epithelial metaplasia, oviductal tumors, polyovular follicles (PF) and polyfollicular ovaries. Female reproductive tracts in mice exposed prenatally to estrogen show altered expression of Hoxa genes and Wnt genes and the analysis of knockout mice lacking Hoxa-10 or Wnt7a show uterine hypoplasia. The growth response of neonatally DES-exposed reproductive organs to estrogen is reduced, as are ER levels and epidermal growth factor (EGF) receptor levels, in addition to other hormone receptor levels. Estrogenic compounds such as bisphenol A (BPA) and nonylphenol as well as dioxins and PCBs were found in the human umbilical cord. BPA can easily cross the placenta and enter the fetus in Japanese monkey and mice. BPA can be found in fetal brain, testis and uterus when given to pregnant mice and monkeys. Neonatal exposure to a high BPA dose induced ovary-independent vaginal changes, PF and infertility lacking corpora lutera. Prenatal exposure to a low BPA dose induced acceleration of vaginal opening in the offspring. Thus, the developing mammal is sensitive to exposure to estrogenic agents. In order to clarify the molecular mechanisms of these effects, we are studying changes in gene expression patterns induced by perinatal exposure to chemicals or estrogen using differential display and DNA microarray techiniques. We have found genes possibly related to the ovary-independent changes by differential display. We also have clustered groups of genes that are responsive to estrogenic stimuli in uterus by using the DNA microarray system. We need to understand the molecular background of the critical period during development, the low dose effect of estrogenic chemicals and the molecular metabolism of hormones and hormone-like agents in animals including humans.
II. Effect of estrogen on amphibian and fishesDuring embryogenesis, exogenous estrogen exposure induces abnormal sex differentiation and the abnormal bone formation in African clawed frog, Xenopus laevi, the cyprinodont fish, mummichog (Fundulus heteroclitus) and mosquitofish (Gambusia affinis). To analyze the function of estrogen, we have isolated cDNA clones of ER a and b from F. heteroclitus, G. affinis and American alligators. The estrogen-responsive genes must play important roles. We try to isolate the estrogen-responsive genes to understand the molecular physiology of estrogen action. Japanese tree frog (Hyla japonica) takes water through ventral skin. We found that sex steroids and endocrine disruptors interfere with water absorption through ventral skin in frogs. Further, using the amphibian and fish as model animals we aim to analyze the effects of numerous chemicals released into the environment on endocrine system function in wildlife. III. Molecular Target SearchAbnormalities caused by endocrine disrupting chemicals are reported but the molecular mechanisms of the effects are not well studied. Although estrogen receptor is one of the strongest candidates possibly responsible for the endocrine disrupting function of many chemicals, it alone cannot explain the variety of phenomena induced by endocrine disrupting chemicals. Thus, we are also looking for new target molecules that may be responsible for endocrine disruption. In parallel, we also are studying the ligand-binding mechanisms of nuclear receptors to hormones and other chemicals using Surface Plasmon Resonance technology. Publication List:Buchanan, D.L., Ohsako, S., Tohyama, C., Cooke, P.S. and Iguchi, T. (2002) 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) inhibition of estrogen-induced mouse uterine epithelial mitogenesis involves changes in cylin and transforming growth factor-beta expression. Toxicol. Sci. 66, 62-68. Honma, S., Suzuki, A., Buchanan, D.L., Katsu, Y., Watanabe, H. and Iguchi, T. (2002) Low dose effect of in utero exposure to bisphenol A and diethylstilbestrol on female mouse reproduction. Reprod. Toxicol. 16, 117-122. Katsu, Y., Takasu, E. and Iguchi, T. (2002) Estrogen-independent expression of neuropsin, a serine protease in the vagina of mice exposed neonatally to diethylstilbestrol. Mol. Cell. Endocr. 195, 99-107. Miyagawa, S., Buchanan, D.L., Sato, T., Ohta, Y., Nishina, Y. and Iguchi, T. (2002) Characterization of diethylstilbestrol-induced hypospadias in female mice. Anat. Rec. 266, 43-50. Okada, A., Ohta, Y., Buchanan, D.L., Sato, T. and Iguchi, T. (2002). Effect of estrogens on ontogenetic expression of progesterone receptor in the female rat reproductive tract. Mol. Cell. Endocr. 195, 55-64. Shimamura, M., Kodaira, K., Hino, K., Ishimoto, Y., Tamura, H. and Iguchi, T. (2002) Comparison of antiandrogenic activities of vinclozolin and D,L-camphorquinone in androgen receptor gene transcription assay in vitro and mouse in utero exposure assay in vivo. Toxicology 174, 97-107. Suzuki, A., Sugihara, A., Uchida, K., Sato, T., Ohta, Y., Katsu, Y., Watanabe, H. and Iguchi, T. (2002) Developmental effects of perinatal exposure to bisphenol-A and diethylstilbestrol on reproductive organs in female mice. Reprod. Toxicol. 16, 107-116. Uchida, K., Suzuki, A., Kobayashi, Y., Buchanan, D.L., Sato, T., Watanabe, H., Katsu, Y., Suzuki, J., Asaoka, K., Mori, C., Arizono, K. and Iguchi, T. (2002). Bisphenol-A administration during pregnancy results in female exposure in mice and monkeys. J. Health Sci. 48, 579-582. Uchida, D., Yamashita, M., Kitano, T. and Iguchi, T. (2002) Oocyte apoptosis during the transition from ovary-like tissue to testes during sex differentiation of juvenile zebrafish. J. Exp. Biol. 205, 711-718. Urushitani, H., Shimizu, A., Katsu, Y. and Iguchi, T. (2002) Early estrogen exposure induces abnormal development of Fundulus heteroclitus. J. Exp. Zool. 293, 693-702. Watanabe, H., Suzuki, A., Mizutani, T., Khono S., Lubahn, D.B., Handa, H. and Iguchi, T. (2002) Genome-wide analysis of changes in early gene expression induced by oestrogen. Genes to Cells 7, 497-507. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |