DIVISION OF MOLECULAR & DEVELOPMENTAL BIOLOGY
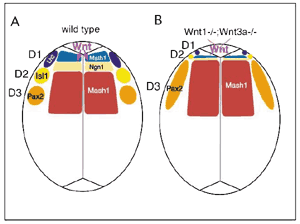
1) Graduate School of Biostudies, Kyoto University The research interest of this laboratory is to understand molecular mechanism of the vertebrate development. Particularly, roles of cell-to-cell signals, including members of Wnt and fibroblast growth factor (FGF) families, are characterized. Evidence indicated that each signal is involved in many aspects of the vertebrate development. For instance, we have revealed that Wnt-3a, a members of Wnt family, plays essential roles in a number of aspects of the mouse development, including somite development, neural crest formation and neural development. However, cellular and molecular mecha-nisms how a cell signaling molecule regulates these different events. Thus, we are focusing on precise functional analysis of cell-to-cell signals and identifica-tion of target genes induced by these signals. In addition, to reveal molecular networks in which these signals are involved, we are also trying genetical approach with the zebrafish. I Roles of Wnt signals during embryogenesisThe Wnt family of genes that encode cysteine rich secreted proteins consists of at least 15 members in the vertabrate. It has already been shown that some of them are expressed and play important roles during gasturulation. For instance, we showed that Wnt-1 and Wnt-3a, which are expressed in the most dorsal region within the developing central nervous system, direct specification of the dorsal interneurons. Analysis of mouse embryos lacking both Wnt1 and Wnt3a and culture of explants from the neural plate indicated that these Wnt signals promote generation of the most dorsal subclass of the interneuron, called D1 and D2, at the expense of that of more ventral subclass, called D3 (FIG. 1). Wnt-3a is also expressed in the primitive streak ectoderm during gastrulation and in the tailbud in later development of the mouse. For dissecion of the complex developmental events regulated by Wnt-3a signaling in these regions, it is important to identify genes regulated by this signal. It has already been demonstrated that T (Brachyury) is a direct target of Wnt-3a in the anterior primitive ectoderm, which is fated to give rise to the paraxial mesoderm, suggesting that Wnt-3a modurates a balance between mesodermal and neural cell fates via T. To gain more insight into roles of Wnt signaling during embryogenesis, we searched for potential target genes of this signaling by an induction gene trap screening in mouse ES cells. In at least three ES cell clones among 794 clones screened, expression of beta-geo reporter genes was dramatically changed in response to the conditioned medium of Wnt-3a expressing cells. The expression analysis of the reporter genes in embryos generated from these ES cell clones revealed that the spatiotemporal expression patterns of these reporter genes were well correlated to those of several Wnt genes. These results suggested that an induction gene trap approach is effective for screening of target genes of Wnt signaling during embryogenesis.
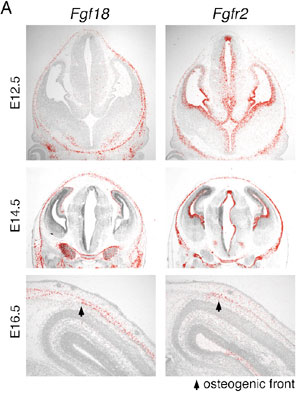
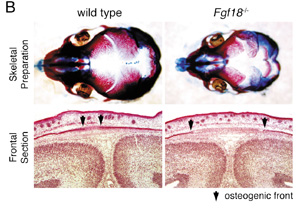
II Roles of Fgf signals in bone developmentThe Fgf family of cell signaling molecules is composed of at least 22 members in the mouse. Although Fgf signaling has been implicated in the bone development, study on null mutant mice have not yet fully shown the role of this family in the skeletal development. As an attempt to identify a candidate Fgf gene essential for bone formation, we have examined the expression of mouse Fgf18 gene using in situ hybridisation and demonstrated expression during calvarial and long bone development. Furthermore, to investigate the role of Fgf18 in vivo, we generated a null allele of the Fgf18 locus by homologous recombination in ES cells. In Fgf18-deficient mouse embryos generated by gene targeting, progress of the suture closure is delayed (FIG. 2). Furthermore, terminal differentiation of the osteoblast is specifically delayed in developing calvaria and long bones. Proliferation of calvarial osteogenic mesenchymal cells is transiently decreased. On the other hand, the number of proliferating and differentiated chondrocytes is increased in the limbs. Fgf18 is thus essential for distinct aspects of cell proliferation and differentiation in bone formation. Thus, Fgf18 plays essential roles in the osteogenesis and the chondrogenesis of the mammal and regulates differently cell proliferation and differentiation in these two processes. To reveal the molecular mechanism that regulates the osteogenesis and the chondrogenesis, we are currently analyzing function of several genes, which act on these processes with Fgf-18.
III Screening of mutations affecting mesoderm development in zebrafishTo identify genes involved in several aspects during early embryogenesis of the vertebrate, we have performed screening of zebrafish mutants induced by ENU, a chemical mutagen. Until now, we have screened 630 of F2 families and found a number of mutants whose phenotypes are different from those already reported. For instance, some of these mutants displayed defects in the somite development. Cloning of genes that are responsible for these defects is in progress. Publication List:Jimbo, T,, Kawasaki, Y,, Koyama, R,, Sato, R,, Takada, S,, Haraguchi, K,, & Akiyama, T. (2002) Identification of a link between the tumour suppressor APC and the kinesin superfamily. Nat. Cell Biol. 4 323-327 Koshida, S., Shinya, M., Nikaido, M., Ueno, N., Schulte-Merker, S., Kuroiwa, A., & Takeda, H. (2002) Inhibition of BMP activity by the FGF signal promotes posterior neural development in zebrafish Dev. Biol. 244, 9-20. Muroyama, Y., Fujihara, M., Ikeya, M,, Kondoh,. H., & Takada, S. (2002) Wnt signaling plays an essential role in neuronal specification of the dorsal spinal cord. Genes Dev. 16, 548-55. Ohbayashi, N., Shibayama, M., Kurotaki, Y., Imanishi, M., Fujimori, T., Itoh, N., & Takada, S. (2002) Fgf18 is required for normal cell proliferation and differentiation during osteogenesis and chondrogeneses. Genes Dev. 16, 870-879. Ueda, Y,, Hijikata, M,, Takagi, S,, Takada, R,, Takada, S,, Chiba, T,, & Shimotohno, K. (2002) Wnt/beta-catenin signaling suppresses apoptosis in low serum medium and induces morphologic change in rodent fibroblasts. Int. J. Cancer. 99, 681-688 Yamanaka, H,, Moriguchi, T,, Masuyama, N,, Kusakabe, M,, Hanafusa, H,, Takada, R,, Takada, S,, & Nishida E. (2002) JNK functions in the non-canonical Wnt pathway to regulate convergent extension movements in vertebrates. EMBO Rep. 3, 69-75 |
||||||||||||||||||||||||||||||||
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |