RESEARCH SUPPORT FACILITY
I. Facilities
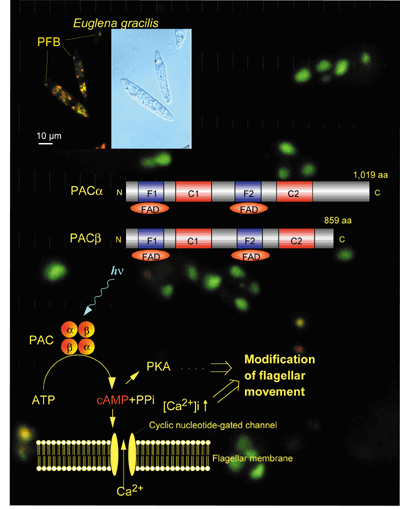
II. Research Activities1. FacultyThe faculty of the Research Support Facility conducts its own research as well as scientific and administrative public services. (1) Photobiology : Photoreceptive and signal transduction mechanisms of phototaxis of unicellular algae are studied action spectroscopically (Watanabe 1995, In “CRC Handbook of Organic Photochemistry and Photobiology”) by measuring computerized-videomicrographs of the motile behavior of the cells at the cellular and subcellular levels. Photoreceptive and signal transduction mechanisms of algal gene expression were also studied by action spectroscopy. A novel blue-light receptor with an effector role was found from Euglena glacilis (Fig. 1; Iseki et al. 2002, Nature 415, 1047-1051): Euglena gracilis, a unicellular flagellate, shows blue-light type photomovements. The action spectra indicate the involvement of flavoproteins as the photoreceptors mediating them. The paraflagellar body (PFB), a swelling near the base of the flagellum has been considered as a photosensing organelle for the photomovements. To identify the photoreceptors in the PFB, we isolated PFBs and purified the flavoproteins therein. The purified flavoprotein (ca. 400 kDa), with noncovalently bound FAD, seemed to be a heterotetramer of a- and b-subunits. Predicted amino acid sequences of each of the subunits were similar to each other and contained two FAD-binding domains each followed by an adenylyl cyclase catalytic domain. The flavoprotein showed an adenylyl cyclase activity, which was elevated by blue-light irradiation. Thus, the flavoprotein (PAC, photoactivated adenylyl cyclase) can directly transduce a light signal into a change in the intracellular cyclic AMP level without any other signal transduction proteins.
(2) Developmental Biology: Replacement of the ankyrin repeats of mouse Notch2 gene with E.coli b-gactosidase gene induces early embryonic lethality around E10.5. The lethality was suggested due to defects in extraembryonic tissues, because the mutant embryo grew and differentiated further in vitro. Histological examination and in situ hybridization analysis with trophoblast subtype-specific probes revealed that the development of giant and spongiotrophoblast cell layers are normal in the mutant placenta, while vasculogenesis in the labyrinth layer apperaed compromised at E9.5. Since the lethality was circumvented by production of chimeric mice with tetraploidy wild type embryos, we concluded that the embryonic lethality is due to defect in growth and/or differentiation of labyrinthine trophoblast cells. The mutant embryo, however, could not be rescued in the tetraploid chimeras beyond E12.5 because of insurfficient development of umbilical cord, indicating another role of Notch2 signaling in the mouse development. Chimeric analysis with diploid wild type, however, revealed contribution of mutant cells to these affected tissues by E13.5. Thus, Notch2 are not cell autonomously required for the early cell fate determination of labyrinthine trophoblast cells and allantoic mesodermal cells, but plays an indispensable role in the further formation of functional labyrinth layer and umbilical cord. (3) Computational Biology: Comparative genomics is a useful approach to find clues to understanding complex and diverse biological systems from rapidly growing genome database. We have constructed a workbench system for comparative analysis of microbial genomes named MBGD. Since the number of completed microbial genome sequences has now become more than a hundred, we have enhanced the efficiency of the database greatly to treat such large number of genomes. One of the unique features of MBGD is to allow users to create their own orthologous classification table with specified set of organisms. By this approach, MBGD accommodates comparative analysis for both closely related and distantly related genomes. We also continue to develop an automated method for orthologous grouping among multiple genomes, which is a key component of MBGD. In addition to splitting fusion genes into orthologous domains, we are also trying to enhance the algorithm to incorporate the information of gene arrangement on each genome. 2. Cooperative Research Program for the Okazaki Large Spectrograph.The NIBB Cooperative Research Program for the Use of the OLS supports about 20 projects every year conducted by visiting scientists including foreign scien-tists as well as those in the Institute. Action spectroscopical studies for various regulatory and damaging actions of light on living organisms, biological molecules, and organic molecules have been conducted (Watanabe, 1995, In "CRC Handbook of Organic Photochemistry and Photobiology". pp, 1276 - 1288). Publication List:I. FacultyIseki, M., Matsunaga, S., Murakami, A., Ohno, K., Shiga, K., Yoshida, K., Sugai, M., Takahashi, T., Hori, T. and Watanabe, M. (2002). A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 415, 1047-1051. Kanesaki, Y., Suzuki, I., Allakhverdiev, Sl., Mikami, K. and Murata, N. (2002) Salt strees and hyperosmotic stress regulate the expression of different sets of genes in Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun. 290, 339-48. Mikami, K., Kanesaki, Y., Suzuki, I. and Murata, N. (2002) The histidine kinase Hik33 perceives osmotic stress and cold stress in Synechocystis sp.PCC 6803. Mol. Microbiol. 46, 905-15. Ohmori, M., Terauchi, K., Okamoto, S. and Watanabe, M. (2002). Regulation of cAMP-mediated photosignaling by a phytochrome in the cyanobacterium Anabaena cylindrica. Photochem. Photobiol. 75, 675-679. Takami, H., Takaki, Y., Uchiyama, I. (2002) Genome sequence of Oceanobacillus iheyensis isolated from the Iheya Ridge and its unexpected adaptive capabilities to extreme environments. Nucleic Acids Res. 30, 3927-3935. Uchiyama, I. (2003) MBGD: microbial genome database for comparative analysis. Nucleic Acids Res. 31, 58-62. II. Cooperative Research Program for the Okazaki Large Spectrograph Iseki, M., Matsunaga, S., Murakami, A., Ohno, K., Shiga, K., Yoshida, K., Sugai, M., Takahashi, T., Hori, T. and Watanabe, M. (2002). A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 415, 1047-1051. Ohmori, M., Terauchi, K., Okamoto, S. and Watanabe, M.(2002). Regulation of cAMP-mediated photosignaling by a phytochrome in the cyanobacterium Anabaena cylindrica. Photochem. Photobiol. 75, 675-679. Toyoshima, M., Takinami, S., Hieda, K., Furusawa, Y. and Negishi, T.(2002).The involvement of cell cycle checkpoint-mutations in the mutagenesis induced in Drosophila by a longer wavelength light band of solar UV. Photochem. Photobiol. Sci., 1, 178 - 183. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |