DIVISION OF CELLULAR REGULATION
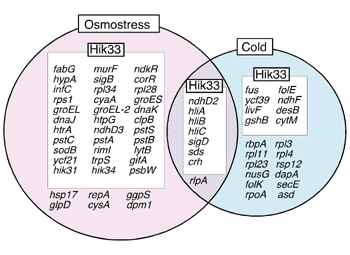
1)from the Biological Research Center, Szeged, Hungary The major thrust of the research efforts of this division is directed towards understanding the signal transduction mechanisms operating in plants and microorganisms with respect to perception of various kinds of stress that arise due to changes in environmental conditions, with particular emphasis on extreme temperatures and salinity. In addition, techniques are being developed for transformation of plants so that they gain the ability to acclimate to and tolerate such stress conditions. In 2002, significant progress was made in the following areas using cyanobacteria as a model system. I. Membrane rigidification enhances the cold inducibility of gene expressionOur previous experiments have indicated that the expression of the cold-inducible desA gene in Synechocystis sp. PCC 6803 (hereafter Synechocystis), which encodes the D12 fatty acid desaturase, is induced by catalytic hydrogenation of fatty acids in plasma-membrane lipids. Thus, we hypothesized that changes in membrane fluidity/rigidity are primary signal of cold stress and are perceived by a cold sensor that is localized at plasma membranes. To evaluate this hypothesis, we examined the effect of genetic rigidification of membrane lipids on the cold inducibility of gene expression using DNA microarrays. We first inactivated the desA and desD genes for D12 and D6 desaturases, respectively, in Synechocystis by targeted mutagenesis. The resultant desA-/desD- cells contained only mono-unsaturated lipid molecules, whereas wild-type cells contained poly-unsaturated lipid molecules. DNA microarray analysis demonstrated that in desA-/desD- mutant cells a large number of genes were induced by cold than in wild-type cells. Fourier transform infrared (FTIR) spectroscopy demonstrated that the replacement of poly-unsaturated membrane lipids by mono-unsaturated ones rigidified the membrane lipids. Thus, we conclude that the rigidification of membrane lipids enhances the cold-inducibility of gene expression. By contrast, the heat inducibility of gene expression was unaffected by mutation of the desA and desD genes, indicating that heat inducibility of gene expression is not triggered by changes in membrane rigidification. These findings suggest that the change in membrane fluidity/rigidity is involved in the mechanism of cold perception but not of heat perception in Synechocystis cells. [Inaba et al. (2003) J. Biol. Chem., in press]. II. Multi-stress sensor Hik33 in SynechocystisAn increase in osmotic pressure leads to a decrease in cell volume (plasmolysis) by efflux of water from the cell. To recover from the plasmolysis, cells perceive a signal of hyperosmotic stress to maintain physiological cellular osmolarity via the expression of osmostress-inducible genes. Screening of Synechocystis mutant library of histidine kinases by DNA microarray technique identified a histidine kinase, Hik33, as an osmosensor. Our previous results have demonstrated that Hik33 is a cold sensor in Synechocystis. DNA microarrays were employed to examine whether Hik33 perceives cold and osmostress differently. The results indicated that Hik33 regulates the expression of a large number of osmostress-inducible genes, although 66% of Hik33-regulated genes encoded proteins of unknown function. Hik33-regulated genes are involved in the structural maintenance of cell wall and membranes and the regulation of phosphate transport, photosynthesis, signal transduction, gene expression, and folding and turnover of proteins (Fig. 1). Although Hik33 regulates the cold-inducible expression of genes which are involved in the regulation of photosynthesis and gene expression, it is clear that Hik33 regulates distinct sets of genes under osmostress and cold conditions with the exception of a small number of genes whose expression was induced by both osmostress and cold (Fig. 1). Therefore, we conclude that Hik33 senses cold and osmostress differently to regulate the expression of distinct sets of genes in a stress-specific manner [Mikami et al. (2002) Mol. Microbiol. 46, 905-915]. Recently, a histidine kinase, NblS, which is a putative homologue of Hik33 in Synechococcus elegatus PCC 7942, was identified as a sensor of strong light and nutrient stress. Moreover, Hik33 was originally identified as DspA, a chemical sensor of drugs such as inhibitors of photosynthesis. These results suggest that Hik33 might also recognize strong light, nutrient stress and chemicals. Confirmation of this possibility is in progress with DNA microarray technique.
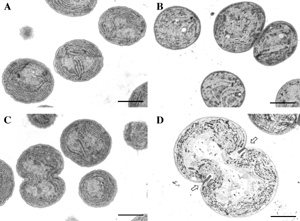
III. Glucosylglycerol, a compatible solute, sustains cell division under salt stress.When the organisms are exposed to a sudden increase in the external concentration of salts, they accumulate compatible solutes, such as trehalose and glycinebetaine, as a result of synthesis de novo or uptake from the environment. Synechocystis cells accumulate glucosylglycerol (GG) and sucrose as the compatible solutes under salt stress. While the molecular mechanism for GG biosynthesis including regulation of the expression of the ggpS gene, which encodes glucosylglycerol phosphate synthase, has been intensively investigated, the role of GG in protection against salt stress remains poorly understood. We studied the role of GG in the tolerance to salt stress, and found that salt stress inhibited cell division and significantly increased cell size in DggpS mutant cells [Ferjani et al., (2003) Plant Physiol. in press; Fig. 2]. Electron microscopy revealed that, in DggpS cells, separation of daughter cells was incomplete and aborted division could be recognized by the presence of a structure that resembled a division ring (Fig. 2). Addition of GG to the culture medium protected DggpS cells against salt stress and reversed the adverse effects of NaCl on cell division and cell size. These observations suggest that GG is important for salt tolerance and thus for the proper division of cells under salt stress conditions.
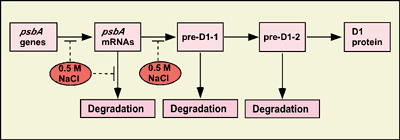
IV. Salt stress inhibits the repair of photosystem II by suppressing the activities of transcriptional and translational activityStrong light impairs the photosynthetic apparatus, in particular that of photosystem II (PSII), via a process known as photodamage or photoinhibition. However, photodamaged PSII can be repaired, and the repair process involves the replacement of the damaged D1 protein (D1), that is a reaction center protein of PSII, by light-dependently synthesized D1 de novo. Under natural conditions light and salt stress are major environmental factors that limit the efficiency of photosynthesis. However, we have found that the effects in vivo of light and salt stress on PSII are completely different in Synechocystis [Allakhverdiev et al. Plant Physiol. (2002) 130, 1443-1453]. Strong light induced photodamage to PSII, whereas salt stress inhibited the repair of the photodamaged PSII and did not accelerate damage to PSII directly. The combination of both stresses inactivated PSII more rapidly as a consequence of their synergistic effects. Radioactive labeling of cells revealed that salt stress inhibited the synthesis of proteins de novo and, in particular, the synthesis of the D1. DNA microarray analysis indicated that the light-induced expression of various genes was suppressed by salt stress. These results suggest that salt stress inhibits the repair of PSII via suppression of the activities of the transcription and translational machinery (Fig. 3).
List of publication:(1) Original articlesAllakhverdiev, S.I, Nishiyama, Y., Miyairi, S., Yamamoto, H., Inagaki, N., Kanesaki, Y., Murata, N. (2002) Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in Synechocystis. Plant Physiol., 130, 1443-1453. Kanesaki, Y., Suzuki, I., Allakhverdiev, S.I., Mikami, K., Murata, N. (2002) Salt stress and hyperosmotic stress regulate the expression of different sets of genes in Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun., 290, 339-348. Mikami, K., Kanesaki, Y., Suzuki, I., Murata, N. (2002) The histidine kinase Hik33 perceives osmotic stress and cold stress in Synechocystis sp. PCC 6803. Mol. Microbiol., 46, 905-915. Mohanty, A., Kathuria, H., Ferjani, A., Sakamoto, A., Mohanty, P., Murata, N., Tyagi, A.K. (2002) Transgenics of an elite indica rice variety Pusa Basmati 1 harbouring the codA gene are highly tolerant to salt stress. Theor. Appl. Genet., 106, 51-57. Simon, W.J., Hall, J.J., Suzuki, I., Murata, N., Slabas, A.R. (2002) Proteomic study of the soluble proteins from the unicellular cyanobacterium Synechocystis sp. PCC6803 using automated matrix-assisted laser desorption/ionization-time of flight peptide mass fingerprinting. Proteomics, 2, 1735-1742. Yamaguchi, K., Suzuki, I., Yamamoto, H., Lyukevich, A., Bodrova, I., Los, D.A., Piven, I., Zinchenko, V., Kanehisa, M., Murata, N. (2002) A two-component Mn2+-sensing system negatively regulates expression of the mntCAB operon in Synechocystis. Plant Cell, 14, 2901-2913. (2) Review articlesChen, T.H.H., Murata, N. (2002) Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol., 5, 250-257. Dmitry, D.A., Murata, N. (2002) Sensing and responses to low temperature in cyanobacteria. In Cells and Molecular Responses to Stress (Storey, K.B. Storey, J.M., eds.), vol. 3, Sensing, Signaling and Cell Adaptation. Elsevier Press, Amsterdam, pp. 139-153 Sakamoto, A., Murata, N. (2002) The role of glycine betaine in the protection of plants from stress: clues from transgenic plants. Plant Cell Environ., 25, 163-171. Sakamoto, T., Murata, N. (2002) Regulation of the desaturation of fatty acids and its role in tolerance to cold and salt stress. Curr. Opin. Microbiol., 5, 208-210. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |