DIVISION OF MOLECULAR NEUROBIOLOGY
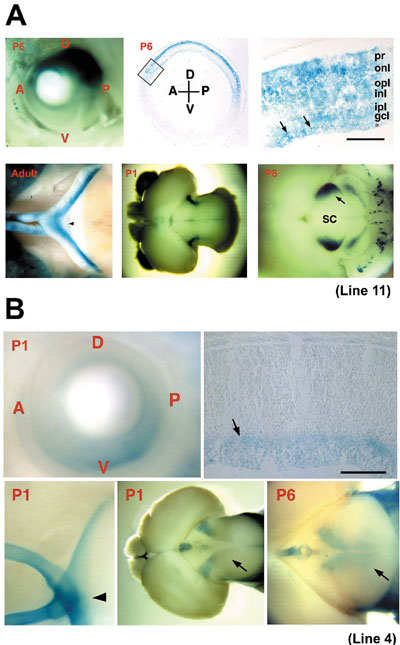
1) CREST, JST. 2) PREST, JST We have been studying the molecular and cellular mechanisms underlying the development of the vertebrate central nervous system. It covers all the major events including the patterning and growth of the nervous system, neuronal determination, axonal navigation and targeting, synapse formation and plasticity, and neuronal regeneration, especially in the visual system. The scope of our interests also encompasses various functions of the matured brain including sensation, behavior, learning and memory. I. Molecular mechanism of neural development (Regional specification in the retina and topographic retinotectal projection)Topographic maps are a fundamental feature of neural networks in the nervous system. Understanding the molecular mechanisms by which topographically ordered neuronal connections are established during development has long been a major challenge in developmental neurobiology. The retinotectal projection of lower vertebrates including birds has been used as a readily accessible model system. In this projection, the temporal (posterior) retina is connected to the rostral (anterior) part of the contralateral optic tectum, the nasal (anterior) retina to the caudal (posterior) tectum, and likewise the dorsal and ventral retina to the ventral and dorsal tectum, respectively. Thus, images received by the retina are precisely projected onto the tectum in a reversed manner. Since 1992, we have been devoting our efforts to searching for molecules with asymmetrical distribution in the embryonic chick retina, and to characterization of their roles in the topographic retinotectal projection. We identified 33 molecules along the nasotemporal axis and 20 molecules along the dorsoventral axis, with various asymmetrical expression patterns in the developing retina. We elucidated the primary structures of all these cDNA clones and examined their expression patterns during development. These included many novel molecules together with the known molecules: transcription factors, receptor proteins, secretory factors, intracellular proteins, and so on. We first identified two winged-helix transcriptional regulators, CBF-1 and CBF-2, expressed in the nasal and temporal retina, respectively. Misexpression experiments using a retroviral vector showed that these two transcription factors determine the regional specificity of the retinal ganglion cells, namely, the directed axonal projections to the appropriate tectal targets along the anteroposterior axis. Secondly, we identified a novel retinoic acid-generating enzyme, RALDH-3, which is specifically expressed in the ventral region of the retina, together with a dorsal-specific enzyme RALDH-1. Furthermore, we recently identified a novel secretory protein, Ventroptin, which has BMP-4 neutralizing activity. Ventroptin is expressed in the retina with a ventral high-dorsal low gradient at early stages. This expression pattern is complementary to that of BMP-4. At later stage (E6~), a nasal high-temporal low gradient expression pattern is also detected. Ventroptin thus shows a double-gradient expression profile along the dorsoventral and nasotemporal axes. Misexpression of Ventroptin altered expression patterns of several topographic genes: BMP-4, Tbx5 and cVax along the dorsoventral axis and ephrin-A2 along the nasotemporal axis. Consistently, in these embryos, projection of the retinal ganglion cell axons to the tectum was also changed along the both axes. The topographic retinotectal projection along the dorsoventral and anteroposterior axes thus appears to be controlled not independently but in a highly concerted manner by Ventroptin. To understand the molecular mechanism underlying the specificity of precise neuronal connections, detailed information on the behavior of developing axons during their pathway selection and target recognition is essential. For such studies, several genetic approaches to the laveling and visualizing of neural projections in mice using transgenic or knock-in techniques have been reported. We employed GAP-lacZ as an axon-targeted reporter protein constructed by fusing the membrane-anchoring domain of the GAP-43 protein to lacZ. The reporter gene was introduced into the genome under the control of a promoter element of Brn3b transcription factor to establish transgenic mouse lines. The individual lines thus generated afforded clear images of specific axonal pathways of the visual (Fig. 1), vomeronasal, pontocerebellar, and auditory systems. The reporter protein labelled the entire axonal process as well as the cell body of developing and mature neurons on staining with X-gal. We showed that these lines facilitate the developmental and anatomical study of these neural systems. Moreover, this strategy must be applicable to a variety of neural systems by using various specific promoter elements. Currently, with respect to the identified topographic molecules, we are conducting misexpression experiments using chick embryos and generating knock-out and transgenic mice to elucidate the molecular functions. We expect that our studies will lead to dissection of the molecular mechanism underlying the retinal patterning and topographic retinotectal projection, and ultimately to uncovering the basic principles for establishing complicated but extremely precise neural networks.
II. Physiological roles of protein tyrosine phosphatase receptor type Z (Ptprz)Protein tyrosine phosphorylation plays crucial roles in various aspects of brain development and brain functions. In 1994, we found that Ptprz/PTPz/RPTPb, a nervous system-rich RPTP, is expressed as a chondroitin sulfate proteoglycan in the brain. The extracellular region of PTPz consists of a carbonic anhydrase-like domain, a fibronectin-type III-like domain and a serine-glycine-rich region, which is considered to be the chondroitin sulfate attachment region. There exist three splice variants of this molecule: A full-length transmembrane form (PTPz-A); a short transmembrane form (PTPz-B); and a soluble form (PTPz-S) which is also known as 6B4 proteoglycan/phosphacan. PTP z is expressed from the early developmental stage to the adulthood in neurons as well as astrocytes. This suggests that this gene plays variegated roles in the brain development and brain function. We found in 1996 that PTPz binds pleiotrophin/HB-GAM and midkine, closely related heparin-binding growth factors which share many biological activities. The chondroitin sulfate portion of PTPz is essential for the high affinity binding (Kd = ~0.25 nM) to these growth factors, and removal of chondroitin sulfate chains results in a marked decrease of binding affinity (Kd = ~13 nM). We further revealed that chondroitin sulfate interacts with Arg78 of midkine. Pleiotrophin and midkine on the substratum stimulated migration of neurons in the glass fiber assay and Boyden chamber cell-migration assay. Experiments using various midkine mutants with various affinities for PTPz indicated that the strength of binding affinities and the neuronal migration-inducing activities are highly correlated. These results suggest that PTPz is involved in migration as a neuronal receptor for pleiotrophin and midkine. To identify the substrate molecules of PTPz we have recently developed the yeast substrate-trapping system. This system is based on the yeast two-hybrid system with two essential modifications: Conditional expression of v-src to tyrosine-phosphorylate the prey proteins and screening using a substrate-trap mutant of PTPz as bait. Using this system, we successfully isolated a number of candidate clones for substrate molecules or interacting molecules. We found that PTPz interacts with PSD-95/SAP90 family members, SAP102, PSD-95/SAP90 and SAP97/hDlg, which are concentrated in the central synapses mediating protein-protein interactions to form large synaptic macromolecular complexes. Here, the C-terminus of PTPz binds to PSD-95/SAP90 proteins through the second PDZ domain. This suggests that PTPz is involved in the regulation of synaptic function. Furthermore, we identified GIT1/Cat-1 as a PTPz substrate. It is known that Cat-1 regulates Pak, a serine threonine kinase which serves as a target for the small GTP-binding proteins, Cdc42 and Rac, and is implicated in a wide range of cellular events including the cell adhesion and cell morphological change. Pleiotrophin, PTPz and GIT1/Cat-1 might regulate the neuronal migration and neurite extension by controlling the Pak signaling pathway. We are continuing efforts to characterize the other candidate clones. In addition, to study the physiological roles of PTPz in vivo, we generated PTPz-deficient mice in which the PTPz gene was replaced with the LacZ gene in 1997. We are currently studying the phenotype of PTPz-deficient mice biochemically, anatomically, physiologically and ethologically, and have already found abnormalities in behavior, learning and memory, etc. Very recently, we found that PTPz is expressed in gastric epithelial cells and functions as the receptor for VacA toxin secreted from Helicobactor pylori. Surprisingly, PTPz-deficient mice are resistant to VacA and do not thereby develop gastric ulcers, in contrast to the wild-type mice. III. Physiological roles of Nax sodium channelSodium-level sensing system in the brain is essential for the regulation of the sodium and water balance in body fluids. Previously, we demonstrated that Nax (also called Nav2/NaG) gene is expressed in the circumventricular organs and Nax-deficient mice ingest salt in excess. In Nax-deficient mice, c-fos expression in the subfornical organ (SFO) and vascular organ of the lamina terminalis (VOLT) was markedly elevated as compared with wild-type mice. Stimulation of the SFO/VOLT of wild-type animals by infusion of a hypertonic sodium solution into intracerebroventricule (ICV) leads to avoidance of salt intake. In contrast, Nax-deficient mice did not show such aversion.
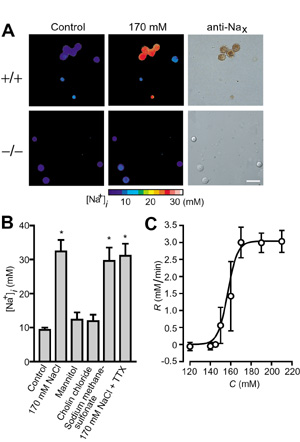
We showed that Nax is a sodium channel which is sensitive to the increase of extracellular sodium level: Entry of sodium ions occurred in the Nax-immunopositive neurons from wild-type mice, in response to a rise of the extracellular sodium concentration (Fig. 2). In contrast, these responses were not observed in Nax-immunonegative cells or neurons of Nax-deficient mutant origin. Transfection of Nax cDNA conferred the sodium sensitivity on Nax-deficient cells. All of the GABA-immunopositive neurons isolated from the SFO responded to the extracellular sodium increase. Based on these findings, we propose that GABAergic inhibitory neurons expressing Nax control the activity of the SFO and suppress the salt-intake behavior of animals under thirst conditions. In addition, we examined the localization of Nax throughout the visceral organs at the cellular level. In visceral organs including lung, heart, intestine, bladder, kidney and tongue, a subset of Schwann cells within the peripheral nerve trunks were highly positive for Nax. An electron microscopic study indicated that these Nax-positive cells were non-myelinating Schwann cells. In the lung, Nax-positive signals were also observed in the alveolar type II cells, which actively absorb sodium and water to aid gas exchange through the alveolar surface. It was thus suggested that the Nax sodium channel is involved in controlling the local extracellular sodium level through sodium absorption activity. Publication List:Hiyama, T.Y., Watanabe, E., Ono, K., Inenaga, K., Tamkun, M.M., Yoshida, S. and Noda, M. (2002) Nax channel involved in CNS sodium-level sensing. Nature Neurosci., 5, 511-512. Watanabe, E., Hiyama, T.Y., Kodama, R. and Noda, M. (2002) Nax sodium channel is expressed non-myelinating Schwann cells and alveolar type II cells in mice. Neurosci. Lett., 330, 109-113. Zubair, M., Watanabe, E., Fukada, M. and Noda, M. (2002) Genetic labelling of specific axonal pathways in the mouse central nervous system. Eur. J. Neurosci., 15, 807-814. Fujikawa, A., Shirasaka, D., Yamamoto, S., Ota, H., Yahiro, K., Fukada, M., Shintani, T., Wada, A., Aoyama, N., Hirayama, T., Fukamachi, H. and Noda, M. (2003) Mice deficient in protein tyrosine phosphatase receptor type Z are resistant to gastric ulcer induction by VacA of Helicobacter pylori. Nature Genet., 33, 375-381 Tanaka, M., Maeda, N., Noda, M. and Marunouchi, T. (2003) A chondroitin sulfate proteoglycan PTPz/RPTPb regulates the morphogenesis of Purkinje cell dendrites in the developing cerebellum. J. Neurosci. (in press) Watanabe, U., Shimura, T., Sako, N., Kitagawa, J., Shingai, T., Watanabe, E., Noda, M. and Yamamoto, T. (2003) A comparison of voluntary salt-intake behavior in Nax-gene deficient and wild-type mice with reference to peripheral taste inputs. Brain Res. (in press) |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |