DIVISION OF DEVELOPMENTAL BIOLOGY (ADJUNCT)
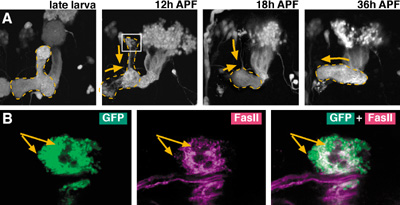
The member of this adjunct division shifted from the Division of Cell Proliferation as of year 2001. The aim of this division is to understand the basic rules by which elaborate neural circuits develop and function. With less than 105 neurons, and subject to powerful molecular and genetic techniques, the brain of the fruit fly Drosophila melanogaster is a good model system for such study. I. Comprehensive identification of neural circuits in the adult brainIn spite of the hundred years of efforts using Golgi and other anatomical techniques, the circuit structure of higher-order associative regions of the brain is still essentially unresolved. The GAL4 enhancer-trap system, which is widely used for mutagenesis and gene cloning of Drosophila, is also a powerful tool for obtaining a vast array of transformant strains that label specific subsets of brain cells. We screened such strains from a stock of 4500 GAL4 lines made by the NP consortium, a joint venture of eight Japanese fly laboratories organized by us. The database of GAL4 expression patterns in the brain of these strains is the infrastructure for further analysis. Having completed the image database of the confocal sections of the adult brain last year, we started making a similar database for the larval brain and accumulated 59,723 brain images so far. Analyses of neurons processing visual and olfactory information continue since previous years. For the visual system, we identified in total 44 pathways that connect the optic lobe and the central brain. To investigate the difference of their roles in visual information processing, we blocked the synaptic transmission of identified neurons with the ectopic expression of the dominant mutant form of Dynamin GTPase, which blocks the recycling of synaptic vesicles. Flies expressing this transgene in various subsets of the 44 visual pathways were subjected to phototaxis assay, which analyzes the flies' ability to identify the orientation of the light source. The effect was different depending on the color of light source, suggesting that different pathways might convey the visual information of different wavelength ranges. For olfactory pathways, last year we identified the structures of the projection neurons that transmit information from the first-order olfactory processing site (antennal lobes) to the second-order sites (mushroom bodies and lateral horns). This year we screened the strains that label neurons of these second-order sites. The area of arborization of these neurons showed strong spatial correlation with the distribution of the terminals of the projection neurons. The olfactory code across glomeruli is thus essentially maintained in the neurons of the second-order sites. Like projection neurons, the spatial specification of these neurons were established before adult eclosion and maintained after surgical ablation of the olfactory input. We also started the characterization of the pathways that convey auditory information. Although the auditory organ of the flies had been identified as the Johnston' Organ, which resides at the base of the antenna, the precise projection patterns of its neurons were not known. We screened the strains that label sensory neurons of this organ and are mapping the distribution of their terminals in the brain. II. Distribution of neurons that release and receive inhibitory transmittersAlthough inhibitory synapses play important roles for information processing, and the GABA is the major neurotransmitter for this function, detail of synaptic connections mediated by GABA is essentially unknown. By combining fluorescent in situ hybridization (FISH) and GFP-immunostaining, we examined the precise distributions of neurons that release and/or receive GABA in the antennal lobe. We found that most of the local interneurons we identified are GABAergic and have GABA receptors. In the case of projection neurons, we found that majority of mACT neurons, but not iACT and oACT neurons, are GABAergic, and that mACT neurons rarely have output sites in the antennal lobe. These findings indicate that, although a wide variety of neurons in the antennal lobe receive inhibitory signals, these signals derive essentially from only the local interneurons. III. Cell lineage-dependent modular structures in the brainThe brain of Drosophila melanogaster is produced by an average of 85 stem cells (neuroblasts) per hemisphere. In previous years, we found that the progeny of each neuroblast project to only a few regions of the adult brain in a stereotypic manner, forming a lineage-dependent circuit module called a clonal unit. In order to map their distribution, we examined the clonal units in more than 800 adult brains and categorized them. We identified 80 candidate units, among which at least 30 have so far been confirmed to be unique. Further characterization and categorization are under way. IV. Cell proliferation in the adult brainRecently, proliferation of neural stem cells in the adult brain was reported in mammals as well as in various insects. To see whether this is also the case in Drosophila, we observed the cell proliferation in the adult brain throughout their lifetime (from zero to 50 days after eclosion). Though we found no systematic proliferation of neuroblasts in old brains, we identified a small number of cells around the antennal nerve that proliferate shortly after eclosion. Immunohistochemistry with a glial cell marker, REPO, and a neuron marker, ELAV, suggests that the proliferating cells are glia. Their progeny becomes either glial cells or the cells of unknown type that is both REPO negative and ELAV negative. V. Development of identified brain structuresThe neural circuits of the adult brain are formed during the larval and pupal stages. Few is known about when and how neurons establish their complicated morphology during pupal metamorphosis. To address this problem, we are tracing the structural change of identified neurons in pupae. An important model circuit for this study is the brain regions called the central complex, whose elaborate structure in the adult brain has long been the scope of anatomical and functional studies. By comparing the image database of the larval and adult brain, we screened for the strains that label putative central complex structure already in larvae. The resulting strains are under detailed investigation to reveal the time course of the central complex composition. VI. Mechanisms underlying the remodeling of neural circuitsLittle is known about the genetic mechanisms that regulate the remodeling of neural circuits during development and maturation of the brain. To address this question, we analyze the larval mushroom body (MB), whose neurons dynamically change the pattern of axon branches during metamorphosis. This process consists of two phases: the degeneration or retraction of existing axon branches, and the re-extension and re-forming of new axon branches (Figure 1A). A single cell labeling of MB neurons revealed that, following the elimination of synapses on the axon branch, axon fibers break and degenerate from proximal to distal subregions of the branches. The synapses lose expression of Fasciclin II (FasII) prior to their elimination (Figure 1B), and the ectopic expression of FasII in the larval MB suppresses the elimination of synapses. Ectopic expression of dominant negative form of Rac, on the other hand, suppresses the degeneration of axon fibers without affecting the synapse elimination. These indicate that the degradation of FasII in the synapses causes the elimination of synapse, whereas the activation of Rac causes the degeneration of axon fibers. IV. Contribution to the science communityAs a joint venture with German and US research groups, we maintain Flybrain, a web-based image database of the Drosophila nervous system (http: //flybrain.nibb.ac.jp). Another database maintained here, mainly for Japanese-speaking fly researchers, is Jfly (http://jfly.nibb.ac.jp). Archives of research-related discussions, experimental protocols, images and movies are provided. Publication List:Hayashi, S., Ito, K., Sado, Y., Taniguchi, M., Akimoto, A., Takeuchi, H., Aigaki, T., Matsuzaki, F., Nakagoshi, H., Tanimura, T., Ueda, R., Uemura, T., Yoshihara, M., and Goto, S. (2002). GETDB, a database compiling expression patterns and molecular locations of a collection of Gal4 enhancer traps. Genesis 34, 58-61. Kido, A., and Ito, K. (2002). Mushroom bodies are not required for courtship behavior by normal and sexually mosaic Drosophila. J Neurobiol 52, 302-11. Kurusu, M., Awasaki, T., Masuda-Nakagawa, L. M., Kawauchi, H., Ito, K., and Furukubo-Tokunaga, K. (2002). Embryonic and larval development of the Drosophila mushroom bodies: concentric layer subdivisions and the role of fasciclin II. Development 129, 409-19. Saito, M, Awasaki, T., and Hama, C. (2002). Genetic analyses of essential genes in cytological region 61D1-2 to 61F1-2 of Drosophila melanogaster. Mol Genet Genomics 268: 446-54
|
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |