DIVISION OF MORPHOGENESIS
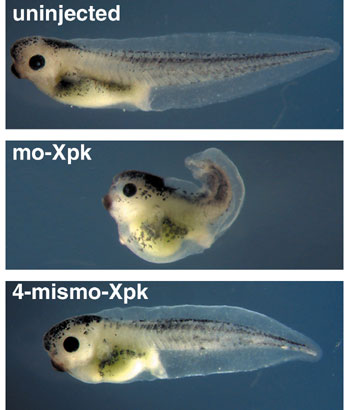
1) Graduate University for Advanced Studies The complex morphogenesis of organisms is achieved by consecutive cell-to-cell interactions during development. Recent studies suggest that growth factors play crucial roles in controlling such intercellular communications in a variety of organisms. In addition to secretory factors, transcription factors which act cell-autonomously are thought to be essential for the determination of cell fates. Our main interest is to know how pattern formation and morphogenesis during development is regulated by these growth factors and transcription factors. We address this problem using several model animals, including frog, fly and acidian, employing embryology, genetics, molecular and cellular biology, and biochemistry. In addition, we have recently introduced array technology to elucidate precise genetic program controlling early development. I. Regulation of growth factor diffusion by proteoglycansDuring early development, cells receive positional information from neighboring cells to form tissue patterns in initially uniform germ layers. Ligands of polypeptide growth factors such as Wnts and TGF-bs are known to participate in this pattern formation as morphogens. As an initial attempt, we searched for potential cell surface proteoglycans which might interact with morphogens and regulate their diffusion. We identified a gene encoding Xenopus glypican-4 (Xgly4), a member of the HSPGs family, and analyzed its role during gastrulation. In situ hybridization revealed that Xgly4 is expressed in the dorsal mesoderm and ectoderm during gastrulation. Xgly4 overexpression and translational inhibition by a Morpholino-oligonucleotide inhibited the gastrulation movements of the embryo and the convergent extension (elongation) of activin-treated animal caps, but did not affect mesoderm induction. Rescue analysis with different mutants of Dsh an essential compomnent of Wnt pathway and Wnt11 demonstrated that Xgly4 function in the non-canonical Wnt pathway, but not in the canonical Wnt/b-catenin pathway for convergent extension movements. Furthermore, we demonstrated that knocking down the levels of Xgly4 inhibits the cell-membrane accumulation of Dishevelled (Dsh), as does the inhibition of Wnt11 in the dorsal marginal zone. Finally, we provided evidence that Xgly4 protein physically binds Wnt ligands. These findings suggest that Xgly4 is serving as a coreceptor for Wnt5A/Wnt11 by presenting these ligands to Frizzled receptor. This hypothesis is further supported by the finding that the causative gene for a zebrafish gastrulation-defective mutant knypek encodes a glypican. II. Gastrulation movement regulated by Wnt signalingGastrulation is one of the most important processes during morphogenesis of early embryo, involving dynamic cell migration and change in embryo shape. Almost all animals undergo gastrulation to form the gut. In spite of its importance, the mechanism underlying the event has just begun to be studied at molecular level. During Xenopus gastrulation, mesodermal cells migrate to the inside of the embryo and move along the blastocoel roof. One of the important mechanisms for this process is convergent extension. As convergent extension begins, cells are polarized and aligned mediolaterally, followed by the intercalation of these cells. As described above, one of the Wnt signaling pathways, called Wnt/JNK (c-Jun N-terminal kinase) pathway, is shown to be important for the regulation of convergent extension. The pathway is highly conserved among species and initially found to be essential for the establishment of planar cell polarity (PCP) of Drosophila wing hair. We demostrated that Xenopus prickle (Xpk), a Xenopus homologue of a Drosophila PCP gene, is an essential component for gastrulation cell movement. Both gain-of-function and loss-of-function of Xpk severely perturbed gastrulation and caused spina bifida embryos without affecting mesodermal differentiation (Figure 1). We also demonstrated that XPK binds to Xenopus Dsh as well as to JNK. This suggests that XPK plays a pivotal role in connecting Dsh function to JNK activation.
We are also studying how Wnt signal is transmitted to Dsh from Frizzled receptors. Protein kinase C (PKC) was implicated in the Wnt pathway, but its molecular role was pooly understood. We searched our Xenopus EST database for novel PKC genes and identified a novel gene encoding PKCd. Loss-of-function of PKCd by a dominant-negative form or Morpholino antisense oligonucleotide has revealed that it is essential for convergent extension during gastrulation. A microscopic analysis showed that PKCd is requierd for the polarization and morphological change of mesodermal cells in this process. This result suggested that PKCd might be involved in the Wnt/JNK pathway. It is known that activation of the Wnt/JNK pathway involves translocation of Dsh from cytoplasm to the membrane and activation of JNK. But the molecular mechanism is not clear. We tested whether PKCd is required for the translocation of Dsh and the activation of JNK, and showed that loss-of-function of PKCd inhibited both of these events. This result indicates that PKCd is essential for Dsh function. We also showed that it forms a complex with Dsh and is translocated to the plasma membrane by Frizzled signaling. In addition, activation of PKCd was sufficient for Dsh translocation and JNK activation. We concluded PKCd plays an essential role in convergent extension by regulating the localization and activity of Dsh in the Wnt/JNK pathway. III. Genetic screening for novel DPP/BMP signaling components utilizing Drosophila model systemDrosophila is one of the ideal model organisms to dissect signal transduction pathway by genetic methods. We have carried out dominant suppressor screening for two transgenic mutant flies expressed constitutively activated DPP/BMP type-I receptors in wing imaginal discs. We isolated 19 suppressor mutants, Suppressor of constitutively activated Dpp singnaling (Scad). Alleles of punt, Mad, shn and dCrebA were found in isolated Scad mutants. We now focus to study a novel locus Scad67. Scad67 encodes a protein with a SP-RING motif. SP-RING motif was originally found in PIAS-family proteins including Drosophila PIAS homolog Su(var)2-10/Zimp. Homozygous Scad67 mutants show embryonic to pupal stage lethality. The most severe zygotic mutant shows abnormal embryonic head structure and segmentation defects. Recent studies have shown that PIAS-family proteins function as a E3-SUMO ligase and enhance SUMO conjugation against specific substrates. Scad67 is an evolutionary conserved molecule and we found two Scad67 homologous genes in human and also in other vertebrates. In the culture cell, transiently expressed vertebrate Scad67s are localized in the nucleus and frequently accumulate in the nuclear dots. Typical PML body markers, such as SP100, CBP, SUMO1 are co-localised with Scad67s. These data indicate that Scad67 facilitates PML nuclear body assemble. This PML assemble activity is clearly enhanced in the presence of SUMO1 and eliminated by inducing point mutation in the Scad67 SP-RING domain. These results strongly suggest that Scad67s are novel family of SUMO-E3 ligases and participate some critical role in the SUMO-conjugation reaction in the nucleus both in vertebrate and invertebrate. IV. Brachyury downstream notochord differentiation in the ascidian embryoAscidians, urochordates, are one of the three chordate groups, and the ascidian tadpole is thought to represent the most simplified and primitive chordate body plan. It contains a notochord, which is a defining characteristic of chordate embryo composed of only 40 cells. To understand the morphogenesis in this simple system, we have focused on a gene, Brachyury, which is known to play an important role in the notochord development. In ascidian, Brachyury is expressed exclusively in the notochord and the misexpression of the Brachyury gene (Ci-Bra) of Ciona intestinalis is sufficient to transform endoderm into notochord. This gene encodes a sequence-specific activator that contains a T-box DNA-binding domain, and in vertebrates, it is initially expressed throughout the presumptive mesoderm and gradually restricted to the developing notochord and tailbud. The phenotype of the Brachyury mutants in mice and zebrafish revealed that this gene is essential for notochord differentiation. Our goal is to elucidate the down stream pathway of this important gene in ascidian in order to set the stage for understanding not only the formation and function of the notochord but how this important structure has evolved. We conducted the subtractive hybridization screens to identify potential Brachyury target genes that are induced upon Ci-Bra overexpression. Out of 501 independent cDNA clones that were induced cDNAs, 38 were specifically expressed in notochord cells. We characterized 20 of them by determining the complete nucleotide sequences and in situ hybridization analyses which show the spatial and temporal expression patterns of the cDNAs. These potential Ci-Bra downstream genes appear to encode a broad spectrum of divergent proteins associated with notochord formation and function.
V. Comprehensive analysis of developmentally regulated genes using macro/microarrayIn order to examine the global expression profile during early development of Xenopus laevis, we have collected massive EST sequences from three normalized cDNA libraries of early gastrula, neurula and tailbud stage. To date, more than 70,000 ESTs were produced and assembled into 12,447 contigs and 15,361 singlets. Homology analysis shows about 40% of known X. laevis proteins are included in these libraries. Moreover, a few thousand novel X. laevis orthologue genes which share high similarities (E<10-10) with those of other animals are also found. EST sequences and annotational information can be accessed through the web at NIBB X. laevis EST database XDB (http://xenopus.nibb.ac.jp/), and cDNAs are available for request. Using these resources, we generated the NIBB 40k cDNA macroarray, and as an initial case to investigate, we conducted a large-scale gene expression screening of Xnr-1 regulated genes. As a result, we have isolated 74 (0.16%) Xnr-1 up-regulated (ratio Publication ListHyodo-Miura, J., Urushiyama, S., Nagai, S., Nishita , M., Ueno, N. and Shibuya, H. (2002) Involvement of NLK and Sox11 in neural induction in Xenopus development. Genes Cells. 7, 487-496. Koshida, S., Shinya, M., Nikaido, M., Ueno, N., Schulte-Merker, S., Kuroiwa, A. and Takeda, H. (2002) Inhibition of BMP activity by the FGF signal promotes posterior neural development in zebrafish. Dev Biol. 244, 19-20. Kurata, T. and Ueno, N. Xenopus Nbx, a novel NK-1 related gene essential for neural crest formation Dev. Biol. (in press) Morita, K., Flemming, A., Sugihara, Y., Mochii, M., Suzuki, Y. Yoshida, S., Wood, B., Kohara, Y., Leroi, A. M. and Ueno N. (2002) A C. elegans TGF-b, DBL-1, controls the expression of LON-1 that regulates polyploidization and body length. EMBO J. 21, 1063-1073. Nakamura, M. Baldwin, D., Hannaford, S. Palka, J. and Montell, C. (2002)DPR, a member of the immunoglobulin superfamily required for the gustatory response to salt. J. Neurosci. 22, 3463-3472. Ohkawara, B., Iemura, S.-I., ten Dijke, P. and Ueno, N. (2002) Action range of BMP is defined by its N-terminal basic amino acid core. Curr. Biol. 12, 205-209. Yuasa, T., Kataoka, H., Kinto, N., Iwamoto, M., Enomoto-Iwamoto, M., Iemura, S. I., Ueno, N., Shibata, Y., Kurosawa, H. and Yamaguchi, A. (2002) Sonic hedgehog is involved in osteoblast differentiation by cooperating with BMP-2 J. Cell. Physiol. 193, 225-232. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |
 2.0), and 47 (0.10%) down-regulated (ratio
2.0), and 47 (0.10%) down-regulated (ratio  0.5) independent EST clones. Whole-mount in situ hybridization was carried out as secondary screening step, showing high ratio of specific gene expression patterns. These clones encode a variety of signal transduction and transcription regulatory components, and also cytoskeletal components, suggesting dynamic cellular changes in response to the Nodal signals. This also proves DNA array to be an effective screening assay for novel genes which function in the early development of Xenopus,utilizing our ESTs.
0.5) independent EST clones. Whole-mount in situ hybridization was carried out as secondary screening step, showing high ratio of specific gene expression patterns. These clones encode a variety of signal transduction and transcription regulatory components, and also cytoskeletal components, suggesting dynamic cellular changes in response to the Nodal signals. This also proves DNA array to be an effective screening assay for novel genes which function in the early development of Xenopus,utilizing our ESTs.