DIVISION OF REPRODUCTIVE BIOLOGY
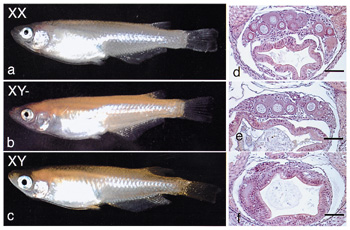
The pituitary-gonadal axis plays an important role in regulating gametogenesis in vertebrates. Gonadotropins typically act through the biosynthesis of gonadal steroid hormones which in turn mediate various stages of gametogenesis. Their effects are particularly profound in teleost fishes which provide several excellent models for investigating the basic hormonal mechanisms regulating gonadal sex differentiation and gametogenesis Our research focuses on (1) the identification of regulators and steroidal mediators involved in sex determination, gonadal sex differentiation and gametogenesis, and (2) the mechanisms of synthesis and action of these mediators. I. Sex-determining gene of medakaAlthough the sex-determining gene SRY/Sry has been identified in mammals, no comparable genes have been found in non-mammalian vertebrates. To identify such a sex-determining gene, a positional cloning approach is suitable. The medaka, Oryzias latipes, has two major advantages for genetic research: a large genetic diversity within the species and the existence of several inbred strains. As in mammals, sex determination in medaka is male heterogametic, although the Y chromosome is not cytogenetically distinct. Alteration of phenotypic sex with no reproductive consequences, and recombination over the entire sex chromosome pair, suggest that there are no major differences, other than a sex-determining gene, between the X and Ychromosomes.To clone positionally the sex-determining region, we generated a Y congenic strain to highlight the genetic differences between the X and Y chromosomesfrom inbred strains of medaka. The Y congenic strain has a sex-determining region derived from the HNI-strain Y chromosome on the genetic background of an Hd-rR strain. Using this strain, we had previously constructed a genetic map of the medaka sex chromosome and constructed a BAC (bacterial artificial chromosome) genomic library. We used recombinant breakpoint analysis to restrict the sex-determining region in medaka to a 530-kb stretch of the Y chromosome. Deletion analysis of the Y chromosome of a congenic XY female further shortened the region to 250-kb (Fig.1). Shotgun sequencing of this region predicted 27 genes. Three of these genes were expressed during sexual differentiation. However, only one gene PG17 was Y specific. The full-length cDNA sequence of PG17 encodes a putative protein of 267 amino acids, including the highly conserved DM domain. The DM domain was originally described as a DNA-binding motif shared between doublesex (dsx) in Drosophila melanogaster and mab-3 in Caenorhabditis elegans. We thus named it DMY (DM-domain gene on the Y chromosome). To establish a role for DMY during sexual differentiation, we screened wild medaka populations for naturally occurring DMY mutants. Two XY females with distinct mutations in DMY were found in separate populations (Awara and Shirone). The first heritable mutant - a single insertion in exon 3 and the subsequent truncation of DMY - resulted in all XY female offspring. Similarly, the second XY mutant female showed reduced DMY expression with a high proportion of XY female offspring. Furthermore, during normal development, DMY is expressed only in somatic cells of XY gonads. These findings strongly suggest that the sex-specific DMY is required for normal testicular development and is a prime candidate for the medaka sex-determining gene.
II. Endocrine regulation of gonadal sex differentiationNile tilapia, Oreochromis niloticus, is an excellent example of the precise nature of steroidogenic actions during gonadal sex differentiation. In this fish, all genetic female (XX) or male (XY) broods can be obtained by artificial fertilization of normal eggs (XX) and sex-reversed, pseudo male sperm (XX), or normal eggs (XX) and super male sperm (YY), respectively. Fertilized eggs hatch after 4 days at 26°C. On the day of hatching, primordial germ cells (PGCs), are located in the outer layer of the lateral plate mesoderm around the hind gut. At 3 days post-hatching, PGCs are located in the gonadal anlagen after the formation of the coelomic cavity in the lateral plate mesoderm rather than through active migration. In tilapia, first morphological sex difference is regarded as the number of germ cells between sexes. Second sexual dimorphism in germ cells is the timing for entry of germ cells into meiosis. Recently, we found that two isoforms of vasa mRNA and protein are present in tilapia. One (vas-s) lacks a part of the N-terminal region found in the other isoform (vas). Both isoforms are expressed in oocytes through the embryonic stage when PGCs localize in the lateral plate mesoderm. Thereafter vas-s expression increases and vas expression becomes undetectable. Expression of both isoforms was observed again after morphological gonadal sex differentiation, irrespective of genotypic sexes. We have also generated medaka transgenic lines with green fluorescent protein (GFP) fluorescence controlled by the regulatory regions of the olvas gene in the germ cells. The intensity of GFP fluorescence increases dramatically in PGCs located in the ventrolateral region of the posterior intestine around stage 25 (the onset of brood circulation). Whole-mount in situ hybridization and monitoring of ectopically located cells by GFP fluorescence suggest that 1) the increase in zygotic olvas expression occurs after PGC specification and 2) PGCs can maintain their cell characteristics ectopically after stages 20-25. The GFP expression persists throughout the later stages in the mature ovary and testis. In tilapia, mitosis of germ cells begins around 10 days post-hatching in genetic females, but can not be confirmed until after sex differentiation in testes of genetic males. Steroid-producing cells in ovaries, but not testes, at the undifferentiated and differentiating stages express all of the steroidogenic enzymes required for estradiol-17 b biosynthesis from cholesterol. Transcripts of estrogen receptors (ER) a and b first appear in both female and male gonads of fry 5-10 days post-hatching. These results, together with evidence of masculinization of genetic females by fadrozole (an aromatase inhibitor) or tamoxifen (an estrogen receptor antagonist), strongly suggest that endogenous estrogens act as the natural inducers of ovarian differentiation in tilapia. In contrast, the ability of steroid-producing cells to synthesize steroid hormones in the testes only appears at the time of testicular differentiation. Transcripts of androgen receptors are not present in gonads of genetic males during sex differentiation. DMRT1 is expressed male-specifically in the gonads (Sertoli cells) during sex differentiation, suggesting an important role of DMRT1 in testicular differentiation. III. Endocrine regulation of spermatogenesisUsing an organ culture system for eel testes consisting of spermatogonia and inactive somatic cells, we have shown that the hormonal regulation of spermatogenesis in eel testes involves the gonadotropin stimulation of Leydig cells to produce 11-ketotestosterone (11-KT), a potent androgen in fish. In turn, 11-KT activates Sertoli cells to stimulate the production of activin B. Addition of recombinant eel activin B to the culture medium induced proliferation of spermatogonia, producing late type B spermatogonia, within 15 days in the same manner as did 11-KT. cDNAs encoding two androgen receptors (AR a and AR b) have been cloned, for the first time in any vertebrates, from eel and tilapia testes. In situ hybridization reveals that although both AR mRNAs are present in eel testes prior to HCG injection, only AR a transcripts increase during HCG-induced spermatogenesis suggesting that AR a and AR b play different roles in spermatogenesis. Activin B binds to activin type I and II receptors on spermatogonia to stimulate de novo synthesis of G1/S cyclins and CDKs leading to the initiation of mitosis. Interestingly, cyclin A1 transcripts are first detected in primary spermatocytes during HCG-induced spermatogenesis in eel testes suggesting an important role for cyclin A1 in the progression to meiosis of male germ cells. IV. Endocrine regulation of oocyte growth and maturationTwo follicular steroidal mediators, estradiol-17b(oocyte growth) and 17a,20b-dihydroxy-4-pregnen-3- one (17a,20b-DP) (oocyte maturation) were identified in several teleost fishes. Two cell-type models in which the thecal layer provides precursor steroids to the granulosa layer, have been demonstrated for estradiol-17b and 17a,20b-DP production. There is a distinct shift in expression of steroidogenic enzyme genes from cytochrome P450 aromatase (P450arom) for estradiol-17b production to 20b-hydroxysteroid dehydrogenase (20b-HSD) for 17a,20b-DP production in granulosa cells immediately prior to oocyte maturation. The preovulatory surge of LH-like gonadotropin is responsible for the rapid expression of 20b-HSD mRNA transcripts in granulosa cells during oocyte maturation. Two possible transcription factors have been identified: Ad4BP/SF-1 (P450arom) and CREB (20b-HSD). Unlike estradiol-17b (genomic action), 17a,20b-DP binds to a novel, G-protein-coupled membrane receptor (non-genomic action), leading to the de novo synthesis of cyclin B, the regulatory component of maturation-promoting factor (MPF), which activates a preexisting 35-kDa cdc2 kinase via phosphorylation of its threonine 161 by a threonine kinase (MO15), thus producing the 34 kDa active cdc2. Upon egg activation, MPF is inactivated by degradation of cyclin B. We showed that the 26S proteasome initiates cyclin B degradation through the first cut of its NH2 terminus at lysine 57. Publication List:Devlin, R.H. and Nagahama, Y. (2002). Sex determination and sex differentiation in fish. Aquaculture 208, 191-366. Hirai, T., Oba, Y. and Nagahama, Y. Fish gonadotropin receptors: molecular characterization and expression during gametogenesis. Fish.Sci. (in press). Huang, Y.S., Yueh, W.S., Huang, J.D., Du, J.L., Sun, L.T., Nagahama, Y. and Chang, C.F. (2002). Cloning and expression of estrogen receptors in the protandrous black porgy (Acanthopagrus schlegeli): Implications of sex change mechanism. Mar. Biotechnol. 4, 236-246. Ikeuchi, T., Todo, T., Kobayashi, T. and Nagahama, Y. (2002). A novel progestogen receptor subtype in the Japanese eel, Anguilla japonica. FEBS Letters 510, 77-82. Kobayashi, T., Kajiura-Kobayashi, H. and Nagahama, Y. (2002). Two isoforms of vasa homologs in a teleost fish: their differential expression during germ cell differentiation. Mech. Develop. 111, 167-171. Kusakabe, M., Kobayashi, T., Todo, T., Lokman, P.M., Nagahama, Y. and Young, G. (2002). Molecular cloning and expression during spermatogenesis of a cDNA encoding testicular 11b-hydroxylase (P45011b) in rainbow trout (Oncorhynchus mykiss).Mol. Reprod. Dev. (in press) Matsuda, M. and Nagahama, Y. (2002). Positional cloning of the sex-determining region of medaka using Y congenic strain. Aquatic Genomics: Steps Toward a Great Future. N. Shimizu, T. Aoki, I. Hirono and F. Takashima (eds.), 236-243, Springer. Matsuda, M., Nagahama, Y., Shinomiya, A., Sato, T., Matsuda, C., Kobayashi, T., Morrey, C.E., Shibata, N., Asakawa, S., Shimizu, N., Hori, H., Hamaguchi, S. and Sakaizumi, M. (2002). DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 417, 559-563. Matsuda, M., Sato, T., Toyazaki, Y., Nagahama, Y., Hamaguchi, S. and Sakaizumi, M. (2002). Oryzias curvinotus has DMY, a gene that is required for male development in the medaka, O. latipes. Zool. Sci. (in press). Morrey, C.E., Nagahama, Y. and Grau, E.G. (2002). Terminal phase males stimulate ovarian function and inhibit sex change in the protogynous wrasse Thalassoma duperrey. Zool. Sci. 19, 103-109. Ohta, K., Rhaman, M.A., Chuda, H., Yoshikuni, M., Nagahama, Y. and Matsuyama, M. Maturation-inducing hormone and it's membrane receptor in gonads of Japanese yellowtail, Seriola quinqueradiata. Fish. Sci. (in press). Rahman, M.A., Ohta, K., Yoshikuni, M., Nagahama, Y., Chuda, H. and Matsuyama, M. (2002). Characterization of ovarian membrane receptor for 17a,20b-Dihydroxy- 4-pregnen-3-one, a maturation-inducing hormone yellowtail, Seriola quinqueradiata. Gen. Comp. Endocrinol. 127, 71-79. Senthilkumaran, B., Sudhakumari, C.C., Chang, X.T., Kobayashi, T., Oba, Y., Guan, G., Yoshiura, Y., Yoshikuni, M. and Nagahama, Y. (2002). Ovarian carbonyl reductase-like 20b-hydroxysteroid dehydrogenase shows distinct surge in messenger RNA expression during natural and gonadotropin-induced meiotic maturation in Nile tilapia. Biol. Reprod. 67, 1080-1086. Tanaka, M., Nakajin, S., Kobayashi, D., Fukada, S., Guan, G., Todo, T., Senthilkumaran, B. and Nagahama, Y. (2002). Teleost ovarian carbonyl reductase-like 20b-hydroxysteroid dehydrogenase: Potential role in the production of maturation-inducing hormone during final oocyte maturation. Biol. Reprod. 66, 1498-1504. Tateno, H., Shibata, Y., Nagahama, Y., Hirai, T., Saneyoshi, M., Ogawa, T., Muramoto, K. and Kamiya, H. (2002). Tissue specific expression of rhamnose-binding lectins in the steelhead trout (Oncorhynchus mykiss). Biosci. Biotechnol. Biochim. 66, 1427-1430. Tokumoto, M., Nagahama, Y. and Tokumoto, T. (2002). A major substrate for MPF: cDNA cloning and expression of polypeptide chain elongation factor 1g from goldfish (Carassius auratus). DNA sequence 13, 27-31. Tokumoto, M., Nagahama, Y. and Tokumoto, T. (2002). Molecular cloning of cDNA encoding a polypeptide chanin elongation factor 1a from goldfish (Carassius auratus). DNA Sequence (in press). Wang, D.S., Kobayashi, T., Senthilkumaran, B., Sakai, F., Sudhakumari, C.C., Suzuki, T., Yoshikuni, M., Matsuda, M., Morohashi, K. and Nagahama, Y. (2002). Molecular cloning of DAX1 and SHP cDNAs and their expression patterns in the Nile tilapia, Oreochromis niloticus. Biochem. Biophys. Res. Comm. 297, 632-640. Yokoi, H., Kobayashi, T., Tanaka, M., Nagahama, Y., Wakamatsu, Y., Takeda, H., Araki, K., Morohashi, K. and Ozato, K. (2002). Sox9 in a teleost fish, medaka (Oryzias latipes): Evidence for diversified function of Sox9 in gonad differentiation. Mol. Reprod. Develop. 63, 5-16. Yoshiura, Y., Senthilkumaran, B., Watanabe, M., Oba, Y., Kobayashi, T. and Nagahama, Y. (2002). Synergistic expression of Ad4BP/SF-1 and cytochrome P-450 aromatase (Ovarian type) in the ovary of Nile tilapia, Oreochromis niloticus, during vitellogenesis suggests transcriptional interaction. Biol. Reprod. (in press). |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |