DIVISION OF GENE EXPRESSION AND REGULATION II
* to Department of Biological Chemistry, University of California-Irvine, California (Feb. 1, 1999- Jan. 15, 2002)
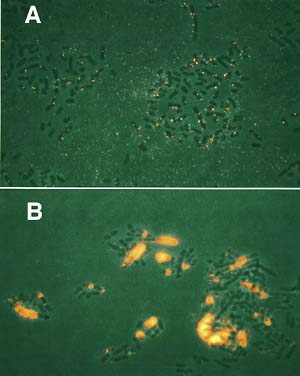
The genomes of higher organisms contain significant amounts of repetitive sequences, which in general, are unstable. At present, neither the physiological function(s) of repeated sequences nor the mechanism controlling the instability is fully understood. To clarify these aspects, we are pursuing the following themes using E. coli, S. cerevisiaeand plants: (1) An amplification mechanism of repeated sequences or genes, especially rRNA repeated genes, (2) a mechanism of replication fork block-dependent recombination, a key reaction that increases or decreases the number of repeats, and (3) development of in vivo artificial gene amplification systems. Functional and structural analyses of the E. coli genome are also being carried out. In 2001, work on the following three subjects has advanced our knowledge of the dynamics and structure of the genome. I. Amplification of Hot DNA segments in Escherichia coliIn yeast, about 150 copies of rDNA are located at a specific locus on chromosome XII. A replication fork-blocking site, called a replication fork barrier (RFB), is located in each rDNA unit. To block the replication fork at RFB, another trans-acting factor, named Fob1 protein, is required. Our recent work indicated that amplification of rDNA repeats requires a DNA replication fork-blocking event. Furthermore, Fob1 protein is essential for rDNA region specific homologous recombination and the production of circular rDNA molecules,as well as rDNA amplification. Such fork blocking dependent recombination was first identified in recombinational hotspot (Hot) in E. coli. The eight kinds of Hot DNAs (HotA-H) were identified using an E. colirnhA- mutant. Among these, enhanced recombination of three kinds of Hot DNAs (HotA, B and C) was dependent on fork blocking events at Ter sites. Then, we examined whether E. coli HotA DNAs are amplified when circular DNA (HotA plus a drug-resistance DNA segment) is inserted into the homologous region on the chromosome of an E. colirnhA- mutant. The resulting HotA DNA transformants were analyzed using pulsed field gel electrophoresis, fluorescence in situ hybridization, and DNA microarrays. The following results were obtained. (1) HotA DNA is amplified by about 40-fold on average. (2) While 90% of the cells contain 6-10 copies of HotA DNA, the remaining 10% of cells have as many as several hundred HotA copies (Fig. 1). (3) Amplification is detected in all other Hot DNAs, among which HotB and HotG DNAs are amplified to the same level as HotA. Furthermore, HotL DNA, which is activated by blocking the clockwise oriC-starting replication fork at the artificially inserted TerL site in the fork-blocked strain with a rnh+ background, is also amplified. II. Yeast RNA polymerase I enhancer is dispensable for both transcription of the chromosomal rDNA gene and cell growth, and its apparent transcription enhancement from ectopic promoters requires Fob1 protein.Previously we developed a system in which various deletions can be introduced into each rDNA repeat in S. cerevisiae. Each rDNA repeat consists of the 35S rRNA gene, the NTS1 spacer, the 5S rRNA gene, and the NTS2 spacer. The FOB1 gene was previously shown to be required for replication fork block (RFB) activity at the RFB site in NTS1, for recombination hot spot (HOT1) activity, and for rDNA amplification and contraction. We have constructed a strain in which the majority of the rDNA repeats are deleted, leaving two copies of rDNA covering the 5S-NTS2-35S region and a single intact NTS1. Growth of the strain is supported by a helper plasmid, carrying, in addition to the 5S rRNA gene, the 35S rRNA coding regions fused to the GAL7 promoter. This strain carries a fob1 mutation, and an extensive expansion of chromosomal rDNA repeats was demonstrated by introducing the missing FOB1 gene by transformation. The budding yeast 35S rDNA transcription enhancer, which is located at the end of the 35S rRNA gene within the rDNA repeats, has been shown to greatly stimulate rDNA transcription in an ectopic reporter system. By removing the single enhancer region remaining in the single NTS1 in the two rDNA copy strain, followed by rDNA amplification, we constructed a yeast strain which was deleted of all of the enhancers from the rDNA repeated genes. We found that this strain did not show any defect in growth or rRNA synthesis. This result suggests that transcription activity measured in the ectopic site is not exactly parallel to that in the rDNA cluster. Ectopic transcription has an effect on recombination. HOT1 DNA enhances recombination at the nearby site when it inserted into a non-rDNA region. HOT1 DNA consist two non-contiguous DNA fragments, called E and I, which are the enhancer and initiator of 35S rDNA transcription, respectively. Thus, it was speculated that HOT1 recombinational enhancement was caused by transcription from the HOT1 region. Later it was proven that a 35S rDNA specific transcription enzyme (PolI) is required for HOT1 recombination. On the other hand, we had previously expected that the fork blocking event is also involved in HOT1 recombination, because the E element contains an RFB site ("enhancer" described above is the same as E except for RFB). Actually, we found that the FOB1 gene is also required for HOT1 activity. However, another group recently reported that HOT1 activity is RFB orientation independent, suggesting that the fork blocking event is not responsible for HOT1 enhancing. We, therefore, examined the effect of the fob1 mutation on HOT1 transcription. The results showed that the mutation abolishes transcription from HOT1 DNA. This means that while Fob1 is a fork blocking protein in the rDNA cluster, it acts as transcription factor as well in the ectopic HOT1 system. Analogously, PolI is essential for HOT1 recombination, but it is never required for homologous recombination in the rDNA cluster. The HOT1 activation mechanism cannot explain any mechanism working in the rDNA cluster. III. E. coli genomic structure and functionWe completed the whole genomic sequence of E. coli W3110, which is a derivative of a common ancestor of MG1655, whose entire genomic sequence was sequenced by the US team. The two strains were separated during or right after World War II. Currently, we are comparing their sequences to understand the micro-evolution of these strains. In order to identify the minimal set of genes required for the duplication of a single cell and to elucidate the function of the genes, we initiated a team project in Japan, headed by Mori (NAIST). For the past two years, our project has been supported by CREST from JST and is being carried out by the following groups: (1) resources, (2) informatics, (3) database, and (4) functional analysis. Until now we have had adequate results, publishing in more than fifty papers and some of the resources established are now available worldwide. In the Annual Bio-training Course held in our Institute this year, our laboratory organized an experimental course entitled "DNA chip: focused on its data analysis" which was strongly supported by team members, belonging to project group (1).
Publication list:Johzuka, K., and Horiuchi, T. (2002) Replication fork-block protein (Fob1) acts as a rDNA region specific recombinator in S. cerevisiae. Genes Cells (in press) Kobayashi, T., Nomura, M., and Horiuchi, T. (2001) Identification of DNA cis-elements essential for expansion of ribosomal DNA repeats in Saccharomyces cerevisiae. Mol. Cell. Biol.21, 136-147. Urawa, H., Hidaka, M., Ishiguro, S., Okada, K., and Horiuchi T. (2001) Enhanced homologous recombination caused by the non-transcribed spacer of the rDNA in Arabidopsis. Mol. Genet. Genomics266, 546-555. Wai, H., Johzuka, K., Vu, L., Eliason, K., Kobayashi, T., Horiuchi, T., and Nomura, M. (2001) Yeast RNA polymerase I enhancer is dispensable for growth and its apparent transcription enhancement for ectopic promoters Fob1 protein implicated in replication and recombination of rDNA. Mol. Cell. Biol. 21, 5541-5553. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |