DIVISION OF CELLULAR REGULATION
1)from the Institute of Basic Biological Problems, Pushchino, Russia
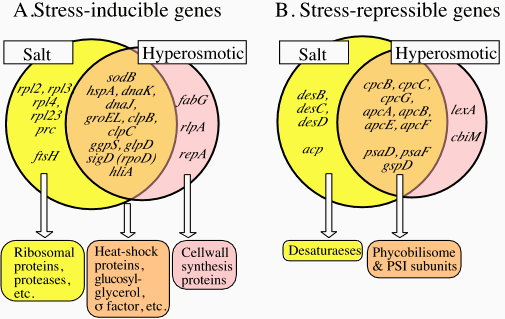
The major thrust of the research efforts of this division is directed towards understanding the signal transduction mechanisms operating in plants and microorganisms with respect to perception of various kinds of stress that arise due to changes in environmental conditions, with particular emphasis on extreme temperatures and salinity. In addition, techniques are being developed for transformation of plants so that they gain the ability to acclimate to and tolerate such stress conditions. In 2001, significant progress was made in the following areas using cyanobacteria as a model system. I. Regulation of gene expression in Synechocystis subjected to salt and hyperosmotic stressSalt stress and hyperosmotic stress are very important environmental factors that severely limit the growth and viability of plants and microorganisms. However, these kinds of stress have been used without proper discrimination in a number of investigations. In this study, these effects were separately investigated on the cytoplasmic volume and gene expression in Synechocystis sp. PCC 6803 (hereafter Synechocystis). Hyperosmotic stress due to 0.5 M sorbitol decreased the cytoplasmic volume by 70% whereas salt stress due to 0.5 M NaCl decreased it by 30% and the effect was only transient. Furthermore, DNA microarray analysis indicated that salt stress strongly induced the expression of genes for some ribosomal proteins and a number of genes for proteins of unknown function. In contrast, hyperosmotic stress strongly induced the expression of the genes for proteins involved in maintenance of cell wall structure (Fig. 1). However, it was also observed that the expression of certain genes was induced by both salt stress and hyperosmotic stress and this category included the genes for heat-shock proteins, the genes for the enzymes that catalyze the synthesis of a compatible solute, glucosylglycerol, and in addition a number of genes for proteins of unknown function (Fig. 1A). Our findings suggest that Synechocystis is capable of discriminating salt stress and hyperosmotic stress as different stimuli [Kanesaki, Suzuki, Allakhverdiev, Mikami and Murata, Biochem. Biophys. Res. Commun. (2002) 290, 339-348]. However, it does not preclude the possibility that mechanisms common to the responses to each form of stress might also exist. Both salt stress and hyperosmotic stress also repressed the expression of certain genes specifically, whereas certain other genes like the genes included in the synthesis of phycobilisome and photosystem I (PSI) subunits were repressed by both (Fig. 1B).
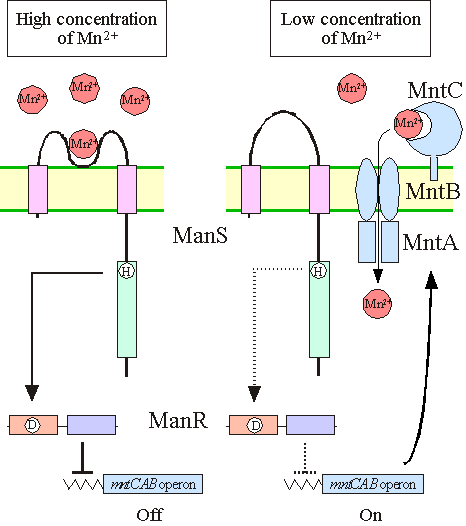
II. Identification of Mn2+ sensor and signal transduction pathwayManganese (Mn) is an essential metal in all organisms. It functions as a cofactor or as a prosthetic group in various enzymes. In particular in the photosynthetic autotrophs which perform oxygenic photosynthesis, four Mn atoms constitute the catalytic center of the oxygen-evolving machinery in a pocket that is formed by the D1 and D2 subunits of photosystem II. Therefore, transport of Mn2+ ions into the cell is crucial for the survival of organisms. It is known that the mntCAB operon that encodes an ABC-type translocater of Mn2+ ions is inducibly expressed in Synechocystis, when the external supply of Mn2+ is limiting. However, the signal transduction mechanism by which cyanobacterial cells recognize the deficiency of Mn2+ ions and induce the downstream expression of the mntCABoperon for the translocater of Mn2+ ions remained to be understood. With this in view a knock-out library of histidine kinases and response regulators of Synechocystis was systematically analyzed by DNA microarray technique. The analysis revealed that mutants of a histidine kinase, ManS, and a response regulator, ManR, exhibited highly induced expression of the mntCAB operon under Mn2+-repleted conditions, suggesting that ManS and ManR constitute the Mn2+-ion sensor and response regulator, respectively, of the signal-transducing pathway in Synechocystis. In the presence of Mn2+, ManS produces a signal to activate ManR and the activated ManR represses the expression of the mntCAB operon, whereas, under Mn2+-depleted conditions, ManS does not generate the signal and therefore ManR exists in its inactive form as a consequence of which the expression of mntCAB operon remains active (Fig. 2). This is the first report on the identification of the functional components of a two-component signal-transduction system by the application of systematic genomics in combination with DNA microarray analysis.
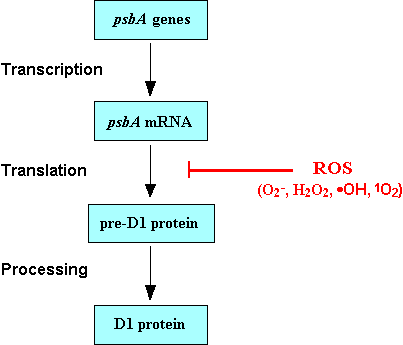
III. Membrane rigidification enhances the cold-induced expression of heat-shock genesChanges in the ambient temperature affect the physical properties of membranes. To obtain insights into the role of membranes in the mechanism of cold signal perception, we have used a mutant of Synechocystis, in which the desA gene for the D12 desaturase and the desD gene for the D6 desaturase are both inactive as a result of targeted mutagenesis. Cells of the desA-desD- mutant synthesizes only a saturated C16 fatty acid and a monounsaturated C18 fatty acid, regardless of the growth temperature, whereas wild-type cells synthesize di-unsaturated and tri-unsaturated C18 fatty acids in addition to the monounsaturated C18 fatty acid [Tasaka et al. (1996) EMBO J. 15, 6416-6425]. In the present study, Fourier transform infrared (FTIR) spectrometry revealed that the desA-desD- mutation rigidified the plasma membrane of Synechocystisat physiological temperatures. We applied DNA microarray technique to examine effects of the membrane rigidification on the induction of gene expression upon cold shock. The results demonstrated that the cold inducibility of certain heat-shock genes, namely, the hspA, htpG and dnaK2 genes, was markedly enhanced by the membrane rigidification. The cold inducibility of cold-shock genes, such as the crh gene for an RNA helicase and the rbp1 gene for an RNA-binding protein, was not further enhanced by the rigidification. Northern blotting confirmed the results of DNA microarray analysis. Our findings suggest that the expression of these heat-shock genes upon cold shock might be regulated by changes in the physical properties of the plasma membrane, supporting the hypothesis that rigidification of membrane lipids is a primary signal for cold sensing. IV. Tolerance of photosynthetic machinery to light stress depends on its ability to repair light-induced photo-damagePhotosynthetic machinery is sensitive to various kinds of environmental stress and, in particular, the photosystem II complex (PSII) is very sensitive to light stress. To overcome this stress photosynthetic organisms have the ability to rapidly repair the PSII from the light-induced damage. We have developed a method to separately measure the damage and repair processes of PSII in Synechocystis, and have examined the effect of various kinds of stress on the damage and repair processes. We have found that the rate of photodamage is proportional to light intensity and this proportionality is unaffected by stress conditions such as oxidative stress, salt stress and cold stress. In contrast, the rate of repair was fast at low intensities of light (and reached the maximum level at relatively low intensities of light), and this rate of repair is depressed by stress conditions. These observations lead us to conclude that photodamage to PSII depends solely on the light intensity, whereas repair of PSII is dependent on the site of regulation by various kinds of environmental stress which are termed "Repair-inhibitory stress". V. Inhibition of the translational machinery by oxidative stressAbsorption of excess light energy by the photosynthetic machinery results in the generation of reactive oxygen species (ROS), such as H2O2, from the photosystem complexes. We investigated the effects in vivo of ROS to clarify the nature of the damage caused by such excess light energy to the photosynthetic machinery in Synechocystis. ROS apparently augments the photodamage to photosystem II (PSII) by inhibiting the repair of the damaged PSII and not by accelerating the photodamage directly. This conclusion was confirmed by the effects of the mutation of genes for H2O2-scavenging enzymes on the repair of PSII. Pulse labelling experiments revealed that ROS inhibited the synthesis of proteins de novo, in particular, that of the D1 protein, a reaction center protein of PSII. Northern blotting revealed that the accumulation of transcripts for the D1 protein was not significantly affected by ROS. Thus, ROS might influence the outcome of photodamage primarily via an effect on translation (Fig. 3).
List of publication:(1) Original articlesAllakhverdiev, S.I., Kinoshita, M., Inaba, M., Suzuki, I., and Murata, N. (2001) Unsaturated fatty acids in membrane lipids protect the photosynthetic machinery against salt-induced damage in Synechococcus. Plant Physiol., 125, 1842-1853. Dilley, R.A., Nishiyama, Y., Gombos, Z., and Murata, N. (2001) Bioenergetics responses of Synechocystis 6803 fatty acid desaturase mutants at low temperature. J. Bioenerg. Biomemb., 33,135-141. Hirani, T.A., Suzuki, I., Murata, N., Hayashi, H., and Eaton-Rye, J.J. (2001) Characterization of a two-component signal transduction system involved in the induction of alkaline phosphatase under phosphate-limiting conditions in Synechocystis sp. PCC 6803. Plant Mol. Biol., 45, 133-144. Inaba, M., Sakamoto, A., and Murata, N. (2001) Functional expression in Escherichia coli of low-affinity and high-affinity Na+/H+ antiporters of Synechocystis. J. Bacteriol., 183, 1376-1384. Nishiyama, Y., Yamamoto, H., Allakhverdiev, S.I., Inaba, M., Yokota, A., and Murata, N. (2001) Oxidative stress inhibits the repair of photodamage to the photosynthetic machinery. EMBO J., 20, 5587-5594. Suzuki, I., Kanesaki, Y., Mikami, K., Kanehisa, M., and Murata, N., (2001) Cold-regulated genes under control of the cold sensor Hik33 in Synechocystis. Mol. Microbiol., 40, 235-244. Turnbull, A.P., Rafferty, J.B., Sedelnikova, S.E., Slabas, A.R., Schierer, T.F., Kroon, J.T.M., Simon, J.W., Fawcett, T., Nishida, I., Murata, N., and Rice, D.W. (2001) Analysis of the structure, substrate specificity, and mechanism of squash glycerol-3-phosphate (1)-acyltransferase. Structure, 9, 1-20. (2) Review articlesSakamoto, A., and Murata, N. (2001) The use of choline oxidase, a glycine betaine-synthesizing enzyme, to create stress resistant transgenic plants. Plant Physiol. (Update), 125, 180-188. Suzuki, I., Los, D.A., and Murata, N. (2001) Perception and transduction of low-temperature signals to induce desaturation of fatty acids. Biochem. Soc. Trans., 28, 628-630. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |