DIVISION OF CELL PROLIFERATION(ADJUNCT)
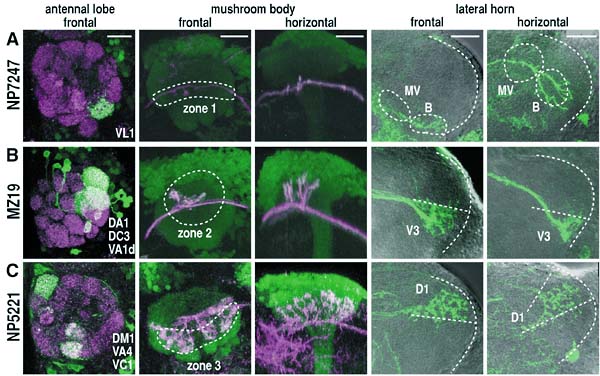
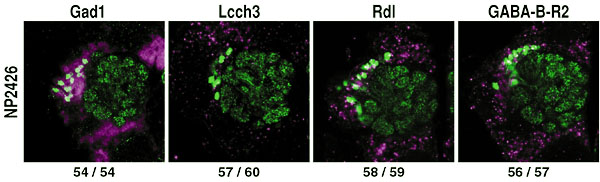
The aim of this adjunct division is to understand the basic rules by which elaborate neural circuits develop and function. With less than 105 neurones, and subject to powerful molecular and genetic techniques, the brain of the fruit fly Drosophila melanogaster is a good model system for investigating the whole of an easily accessible nervous system that shares certain of the architectural and functional features of the more complex vertebrate brains. I. Comprehensive identification of cells in the adult brainIn spite of the hundred years of efforts using Golgi and other anatomical techniques, the circuit structure of higher-order associative regions of the brain is still essentially unresolved. The GAL4 enhancer-trap system, which is widely used for mutagenesis and gene cloning of Drosophila, is also a powerful tool for obtaining a vast array of transformant strains that label specific subsets of brain cells. We screened such strains from a stock of 4500 GAL4 lines made by the NP consortium, a joint venture of eight Japanese fly laboratories organised by us. In the first step, all the lines were crossed with the flies carrying the UAS-GFP transgene, which fluoresces only in GAL4-expressing cells. Their patterns were recorded from freshly dissected, unfixed adult brains with a high-speed confocal microscope. In June 2001, we finished making the image database of 131,406 photographs depicting the confocal sections of 3,939 GAL4 strains. From July 2001, we started making a similar database of the late larval brain using the same set of GAL4 strains. In the second step, useful lines were selected from the database, and fixed and clealised brains were subjected to more precise serial sectioning with conventional confocal microscopes and to three-dimensional reconstruction with UNIX workstations. Although our long-term aim is to get the comprehensive overview of the fly brain neural network, at the initial stage a few brain regions are chosen for intensive study. The first target is to identify projection interneurones that connect lower-level sensory processing sites and higher-order associative regions. These fibres convey olfactory, gustatory, auditory and visual sensory information. For visual pathways, we identified another 10 types of interneurones that project from the optic lobe to the central brain. The total number of identified neurone types of this kind now became 40. Their structures, as well as their role in the controlled movement of flies, are under investigation. For olfactory pathways, we selected 34 strains that label various subsets of antennal lobe projection neurones, which transmit information from the first-order olfactory processing site (antennal lobes) to the second-order sites (mushroom bodies and lateral horns). We found that the terminals of these PNs form clear zonal separation in both second-order sites. We identified three zones in the mushroom bodies and nine zones in the lateral horns (Fig. 1). Projection neurones from several glomeruli, each deriving from a single glomerulus of the antennal lobe, converge at selective zones. Another type of neurones, each contributing multiple glomeruli, terminate at zones that overlap with the zones of uniglomerular neurones. The olfactory code across glomeruli is thus both distributed to, and convergent at, discrete zones of the second-order sites. These stereotypic projection patterns were established before adult eclosion, and surgical ablation of olfactory sense organ caused no significant reorganisation. These suggest that distinct odortypic pathways are established and maintained genetically rather than in an activity dependent manner. II. Mapping of neurotransmitters and receptors in the adult brainTo understand the function of the identified neural network, it is also important to get the information about the types of neurotransmitters and receptors used in these cells. Previously, such information is obtained by staining brain tissue with various antibodies. This approach, however, has the limitation due to the availability of good antibodies. Taking the advantage of the completion of Drosophilawhole genome sequence, we employed a novel approach. From the database, we first search genes that code receptors and enzymes associated to transmitter synthesis. Cells that express those genes are labelled by using in-situ RNA hybridisation. We then screen GAL4 enhancer-trap strains that label cells in the corresponding area, and perform double labelling to certify the colocalisation of GAL4 expression and in-situ label. Second year of this project, we concentrate on GABA, a major inhibitory transmitter, and are mapping the GABA-generating cells and cells that express three types of GABA receptors (Fig. 2). III. Analysis of the cell lineage-dependent modular structures in the brainThe central brain of Drosophila melanogaster is produced by an average of 85 stem cells (neuroblasts) per hemisphere. We visualised the innervation patterns of the progeny of single neuroblasts in the adult brain using the FRT-GAL4 system. In most cases, cell bodies form a tightly packed cluster and their neurites fasciculate to form a single bundle to innervate a limited number of brain regions in a stereotypic manner. These suggest that the progeny of a single neuroblast often form a lineage-dependent circuit module, which we named a "clonal unit." To understand the mechanisms underlying this clonal clustering and fasciculation, we focused on the role of neural-specific homophilic cell adhesion molecules in the cell body layer (cortex) of the developing larval brain. DN-cadherin and Neuroglian are distributed uniformly along the border between all the neurones. FasciclinII (FasII), on the other hand, localises in several clusters of neurones, each of which looks like clonally related. Double labelling of FRT-GAL4 clones and FasII-expressing cells revealed that FasII clusters indeed correspond to clones. The distribution of FasII is limited to the cell border inside the clones. Cell surface flanking the neighbouring clones is free of FasII. Such localisation might infer that FasII would mediate cell-cell adhesion within clonal cluster. fasII mutant clones, however, showed no remarkable defect on the formation of clonal cell clustering. Pan-neuronal ectopic expression of fasII caused little effect, either. The ectopically expressed FasII showed the same characteristic localisation pattern: it concentrates along the intraclonal cell borders but not along the interclonal cell borders. Why doesn't ectopic FasII exist at the interclonal cell borders? One possible explanation is that there might be physical boundary that prevents direct contact of neurones between different clones. We thus examined the arrangement of glial cells in the larval brain. Double labelling of glial cells and FasII-expressing clones showed that a type of glia send extensive processes between neurones. In the outer area of the cell body layer, which is near the brain surface and houses neuroblasts and newly-generated cells, glial processes wrap only the outer surface of the clonal clusters. Processes are not observed within the cluster. Glial cells thus physically separate the clonal border in this area. Deeper in the cell body layer, which consists of old cells, thin glial processes penetrate the boundary between essentially all the neurones. IV. Contribution to the science communityAs a joint venture with German and US research groups, we maintain Flybrain, a web-based image database of the Drosophila nervous system (http://flybrain.nibb.ac.jp). Another database maintained here, mainly for Japanese-speaking fly esearchers, is Jfly(http://jfly.nibb.ac.jp). Archives of research-related discussions, experimental protocols, images and movies are provided. Publication List:Awasaki, T. and Kimura, K. (2001) Multiple function of poxn gene in larval PNS development and in adult appendage formation of Drosophila, Dev. Genes. Evol.211: 20-29 Kurusu, M., Awasaki, T., Masuda-Nakagawa, L. M., Kawauchi, H., Ito, K. and Furukubo-Tokunaga, K. (2002) Embryonic and larval development of the Drosophila mushroom bodies: Concentric layer subdivisions and the importance of fasciclin II. Development, in press Verkhusha, V. V., Otsuna, H., Awasaki, T., Oda, H., Tsukita, S., and Ito, K. (2001) An enhanced mutant of red-fluorescent protein DsRed for double labelling and developmental timer of neural fiber bundle formation. J Biol Chem 276 (32): 29621-29624.
|
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |