DIVISION OF BIOENERGETICS
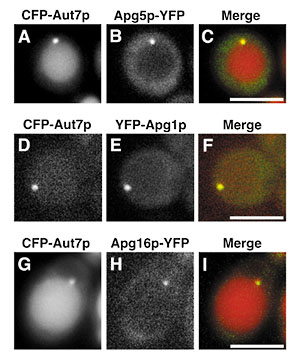
1) PRESTO, JST, Oct 1999-. This division aims to understand the autophagy in respects to its molecular mechanism and its physiological role in higher eukaryotes. Cells execute degradation processes of their constituents together with biosynthetic processes. These two processes are well coordinated to regulate the biological activities. In other word, we must shed light on degradation process to fully understand the cell, because the study on the degradation has been retarded compared to the biosynthetic process. Autophagy is well conserved in eukaryotes and is a major route for bulk degradation of cytoplasmic constituents and organelles in a lytic compartment, lysosome/vacuole. I. BackgroundUpon nutrient starvation, autophagic process starts as building up a membrane structure, an autophagosome, in the cytoplasm. The autophagosome sequesters cytosol and organelles nonselectively. Then it is delivered to the vacuole/lysosome, and the cytoplasmic materials inside are degraded by vacuolar/lysosomal proteases. We had discovered autophagy in a simple eukaryotic model organism, Saccharomyces cerevisiaeand morphologically defined the whole process. We have isolated a set of autophagy-deficient mutants (apg), and have cloned most of the APG genes essential for autophagy. We are now characterizing these gene products and their homologues in mammals and higher plants. 2. Discovery of novel preautophagosomal structureWe have cloned the last APG genes, APG2, and characterized of the gene product. We drew a rough functional map of each Apg proteins this year. This is based on the finding of novel structure called preautophagosomal structure (PAS). Many Apg proteins (Apg1, Apg2, Apg5, Apg8, Apg16) are colocalized on preautphagosomal structure. Autophagy-specific phosphatidylinositol 3-kanise complex (Apg6, Apg14, Vps34 and Vps15) are essential for organization of PAS. Apg8, ubiqitin-like protein, gathers to PAS depending on lipidation reaction and the Apg5-Apg12 conjugates. Apg5 is recruited to PAS depending on Apg16 but independent on Apg12-conjugation. Apg2 is localized on PAS via Apg1. Apg1 protein kinase function in transition from PAS to autophagosome. Thus, PAS is the organizing center of autophagosome.
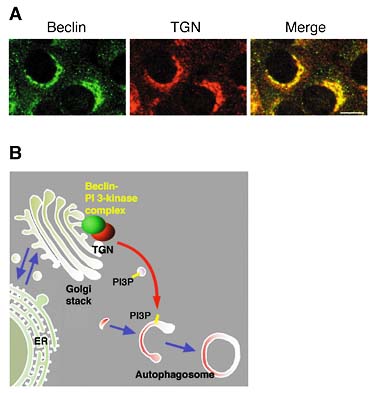
3. Autophagosome formation depends on specific sets of coat proteinsWe revealed that autophagosme fuses with the vacuole by NSF/SNARE meditated mechanism, which is common through endmembrane system, such as ER and Golgi. However, autophagosome formation is not dependent of NSF. Instead, the involvement the proteins of early secretory pathway are revealed. Sec12, Sec16, Sec23, Sec24 are required for autophagosome formation. Interestingly, they are not prerequisite for Cvt pathway, the similar pathway to autophagy. The findings may imply that early phase of autophagosome formation is related to secretory pathway in some way. 4. Molecular dissection of autophagosome formationApg12 and Apg5 are covalently attached in a manner similar to the ubiquitin conjugation system. The Apg12-Apg5 conjugation system is well conserved in mammalian cells. Part of the mammalian Apg12-Apg5 conjugate localized to the isolation membranes when autophagy proceeds, whereas most of it existed in the cytoplasm. Using GFP-tagged Apg5, we revealed that the cup-shaped isolation membrane is developed from a small crescent-shaped compartment. Apg5 localized on the isolation membrane throughout its elongation process. Apg5 was preferentially distributed in the outer side of the membrane and detaches from it immediately before or after autophagosome formation is completed. In contrast, LC3, the mammalian homologue of Apg8, was associated with both sides of isolation membranes and autophagosomes as well as their precursors. To examine the role of Apg5, we generated Apg5-deficient ES cells. APG5-/-cells are viable but bulk protein degradation was significantly reduced. Autophagosome formation was impaired in these cells. The mutant Apg5K130R, which is unable to be conjugated with Apg12, could bind to the autophagosome precursors but could not start elongation of the membranes. Thus, the covalent modification of Apg5 with Apg12 was not required for its membrane targeting but is essential for involvement of Apg5 in elongation of the isolation membranes. Intriguingly, Apg12-Apg5 was required for processing of LC3 and its targeting to the membranes. Therefore, the Apg12-Apg5 conjugate plays essential roles in isolation membrane development in co-operation with LC3. In addition, our studies provided good molecular markers, LC3 and Apg12-Apg5, for autophagic membrane at all stages and isolation membranes, respectively, which so far have been defined only by morphology. 5. Production of phosphatidylinositol 3-phosphate (PI3P) at the trans-Golgi network (TGN) is required for autophagyThis year, we reported that, in yeast, Vps34 PI3-kinase and Apg6 form a protein complex that plays a essential role in autophagy by producing PI3P. Then, we found in mammalian cells that the complex of the mammalian Apg6 homologue, beclin and PI3-kinase was distributed in the TGN rather than autophagosomes (Figure 2A). We propose that beclin functions as a regulatory-subunit of the PI3-kinase complex at the TGN, which supplies PI3P for autophagosome formation (Figure 2B).
Original papers :Ishihara, T., Hamasaki, M., Yokota, S., Suzuki, K., Kamada, Y., Kihara, A., Yoshimori, T., Noda, T., and Ohsumi, Y. (2001) Autophagosome requires specific early Sec proteins for its formation and NSF/SNARE for its fusion to the vacuole. Mol. Biol. Cell12, 3690-3702. Kim, J., Kamada, Y., Stromhaug, P. E. Guan, J., Hefner-Gravinnk, A., Bevan, A., Scott, S. V., Ohsumi, Y., Dunn, Jr., W. A., Klionsky, D. J. (2001) Cvt9/Gsa9 function in sequestering selective cytosolic cargo destined for the vacuole. J. Cell. Biol. 153, 381-396. Kihara, A., Noda, T., Ishihara, N. and Ohsumi, Y. Two distinct Vps34 Ptdlns 3-kinase complex functionin autophagy and CPY. (2001) sorting in Saccharomyces cevisiae. J. Cell. Biol. 152, 519-530. Kihara, A., Kabeya, Y., Ohsumi, Y., and Yoshimori, T. (2001) Beclin-phosphatidylinositol 3-kinase complex functions at the trans-Golgi Network. EMBO Report, 2, 330-335. Komtatsu, M., Tanida, I., Ueno, T., Ohsumi, M., Ohsumi, Y., and Kominami, E. (2001) The C-terminal region of an Apg7p/Ctv2p is required for homodimerization and is essential for its E1-activity and E1-E2 complex-formation. E. J. Biol. Chem.276, 9846-9854. Mizushima, N., Yamamoto, A., Hatano, M., Kobayashi, Y., Kabeya, Y., Suzuki, K., Tokuhisa, T., Ohsumi, Y and Yoshimori, T. (2001) Dissection of autophagosome formation using Apg 5-deficient mouse embryonic stem cells. J. Cell. Biol. 152, 657-667. Shintani, T., Suzuki,K., Kamada, Y., Noda, T., Ohsumi, Y. (2001)Apg2p functions in autophagosome formation on the perivacuolar structure. J. Biol. Chem.276, 30452-30460. Suzuki, K., Kirisako, T., Kamada, Y., Mizushima, N., Noda, T. and Ohsumi, Y. (2001) The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO. J. 20, 5971-5981. Review :Ohsumi, Y. (2001) Molecular dissection of autophagy: Two ubiquitin-lke systems. Nature Reviews, Molecular Cell Biol, 2, 211-216. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |