The Technology Department is a supporting organization for researchers
and research organi-zation within the NIBB. The Department develops
and promotes the institute's research activities and at the same
time, maintains the research functions of the institute.
The Department is organized into two groups: one, the Common Facility
Group, which supports and maintains the institute's common research
facilities and the other, the Research Support Group, which assists
the research activities as described in individual reports.
Technical staffs participate, through the depart-ment, in mutual
enlightenment and education, and increase their capability in
technical area. Each technical staff is proceeded to the fixed
division usually and they support the various research with their
special biological and biophysical techniques.
The Department hosts an annual meeting for technical engineers
who work in various fields of biology at universities and research
institutes throughout Japan. At this meeting, the participants
present their own activities and discuss technical problems. The
Proceedings are published soon after the meeting.
Head of Facility: Mikio Nishimura
Associate Professor: Masakatsu Watanabe
Research Associates:Yoshio Hamada, (Tissue and Cell Culture),
Kenta Nakai (Computer ;-April 30, 1995)
Technical Staff: Mamoru Kubota, Chieko Nanba, Toshiki Ohkawa,
Kaoru Sawada, Tomoki Miwa, Hideko Nonaka, Kimiko Yamamiya (-March
31, 1995), Masayo Iwaki (June 1, 1995-)
The Facility provides large-and medium-scale instruments and facilities
for biophysical, molecular biological, and computational analyses
as well as for growing and maintaining biological specimens. The
facility is shared among the research members, and has seven laboratories,
among which the Large Spectrograph Laboratory and the Laboratory
of Stress-Resistant Plants are dedicated to cooperative use under
the NIBB Cooperative Research Programs.
1. The Large Spectrograph Laboratory
This laboratory provides, for cooperative use, the Okazaki Large
Spectrograph (OLS), which is the largest spectrograph in the world,
dedicated to action spectroscopical studies of various light-controlled
biological processes. The spectrograph runs on a 30 kW-Xenon arc
lamp and has a compound grating composed of 36 smaller individual
gratings. It projects a spectrum of a wavelength range from 250
nm (ultraviolet) to 1,000 nm (infrared) onto its focal curve of
10 m in length. The fluence rate (intensity) of the monochromatic
light at each wavelength is more than twice as much as that of
the corresponding monochromatic component of tropial sunlight
at noon (Watanabe et al., 1982, Photochem. Photobiol.,
36, 491-498).
A tunable two-wavelength CW laser irradiation system is also available
as a complementary light source to OLS to be used in irradiation
experiments which specifically require ultra-high fluence rates
as well as ultra-high spectral-, time-and spatial-resolutions.
It is composed of a high-power Ar-ion laser (Coherent, Innova
20) (336.6-528.7 nm, 20 W output), two CW dye lasers (Coherent,
CR599-01) (420-930 nm, 250-1000 mW output),
A/O modulators (up to 40 MHz) to chop the laser beam, a beam expander,
and a tracking microbeam irradiator (up to 200 Mu ms-1
in tracking speed, down to 2Mum in beam diameter) with an infrared
phase-contrast observation system.
2. Tissue and Cell Culture Laboratory
Various equipments for tissue and cell culture are provided. This laboratory is equipped with safely rooms which satisfy the P2/P3 physical containment level. This facility is routinely used for DNA recombination experiments.
3. Computer Laboratory
To meet various computational needs and to provide means of electronic communication in this Institute, many kind of computers are equipped: VAX/VMS machines, UNIX engineering workstations (SPARCstations, NEWS machines, DECstations), and some personal computers (Macintosh's and Windows machines). All of these machines are connected each other through the Ethernet, the Fast Ethernet, or the CDDI, which are also linked by optical fibers to the high performance multimedia backbone network of Okazaki National Research Institutes. Since this backbone network, called ORION, is joined to the Internet, the users of these machines can access various services and databases on the Internet. Each laboratory has several computers that are also connected to the network of the Institute. The Computer Laboratory provides various computational services to the Institute members: file servers for Macintosh and NetWare users, some print servers that accept printing requests from PC and UNIX users, a computational server that provides sequence analyses and database retrievals, communication servers to the Internet, and so on. The laboratory also provides an information dispatching service to the Internet using the World Wide Web (URL is http://www.nibb.ac.jp).
4. Plant Culture Laboratory
There are a large number of culture boxes, and a limited number of rooms with environmental control for plant culture. In some of these facilities and rooms, experiments can be carried out at the P1 physical containment level.
5. Experimental Farm
This laboratory consists of two 20 m2 glass-houses with precise temperature and humidity control, a small farm, two greenhouses (45 and 88 m2 ) with automatic sprinklers, two open aquariums (30 and 50 t) and several smaller tanks. The laboratory also includes a building with office, storage and work-space.
6. Plant Cell Laboratory
Autotrophic and heterotrophic culture devices and are equipped for experimental cultures of plant and microbial cells. A facility for preparation of plant cell cultures including an aseptic room with clean-benches, is also provided.
7. Laboratory of Stress-Resistant Plants
This laboratory was founded to study transgenic plants with respect
to tolerance toward various environmental stresses. It is located
in the Agricultural Experimental Station of Nagoya University
(30 km from the National Institute for Basic Biology). The laboratory
provides a variety of growth chambers that precisely control the
conditions of plant growth and facilities for molecular biological,
and physiological evaluations of transgenic plants. The laboratory
is also a base of domestic and international collaborations devoted
to the topic of stress-resistant transgenic plants.
1. Faculty
The faculty of the Research Support Facility conducts its own research as well as dcientific and administrative public services.
(1) Photobiology: Photoreceptive and signal trans-duction mechanisms of phototaxis of single-celled, flagellate algae are studied action spectroscopically (Watanabe 1995, In CRC Handbook of Organic Photochemistry and Photobiology) by measuring computerized-videomiceographs of the motile behavior of the cells at the cellular and subcellular levels (Erata et al. 1995, Protoplasma). Photo-receptive and signal tranduction mechanisms of algal gametogenesis are also studied by action spectroscopy.
(2) Developmental Biology: Notch is an integral cell surface membrane protein that is known to play a key role in developmental cell-cell interactions in Drosophila, particularly in lateral specification of neural versus epidermal cell fates, a process described thus far only in invertebrates. It is thought to act by a direct signaling pathway rather than through one of the classical signal transduction cascades. The mammalian genome is known to contain three Notch homologues but their developmental significance is not clear. To investigate their role in mammalian development, we have sequenced the murine Notch 2 cDNA, determined the primary sequence of its protein, and have investigated its genomic organization. We are now attempting to produce a mutant in which the ankyrin repeat region of Notch 2 is replaced by lacZ. Analysis of the mutant phenotype will provide us with insights about the significance of the repeat in Notch 2 signal transduction in relation to its developmental importance.
(3) Computational Biology: Efforts to develop new methodology
for sequence analysis have been continued. A system to predict
protein localization sites from the sequences had been developed
and is now open to the public through the Internet (http://psort.nibb.ac.jp).
Sequence motifs are thought to be good indices of biological functions.
Methodology of protein sequence analysis based on sequence motifs
has been studied and is implemented as a computer program. The
product of computer assistance for sequence analysis is being
constructed using the World Wide Web.
Many kind of databases become available in recent years, but almost
all of them are those of molecular information such as nucleic
acids or amino acids sequences. It is also desired to construct
a database for biological functions like networks of the regulation
of gene expression. As a model case, Bacillus subtilis genome
database is constructed to represent the categorized classification
of gene products in collaboration with Dr. N. Ogasawara (NAIST)
and Human Genome Center, University of Tokyo. Other kind of biological
functional databases including image data are planned to represent
the accumulated knowledge of development, differ-entiation, and
morphogenesis.
2. Cooperative Research Program for the Okazaki Large Spectrograph
The NIBB Cooperative Research Program for the Use of the OLS supports
about 30 projects every year conducted by visiting scientists
including foreign scientists as well as those in the Institute.
Action spectroscopical studies for various regulatory and damaging
actions of light on living organisms, biological molecules, and
organic molecules have been conducted (Watanabe, 1995, In
CRC Handbook of Organic Photochemistry and Photobiology).
I. Faculty
Erata, M., Kubota, M., Takahashi, T., Inouye, I. and Watanabe, M. (1995) Ultrastructure and phototactic action spectra of two genera of cryptophyte flagellate algae, Cryptomonas and Chroomonas. Protoplasma, 188, 258-266.
Higuchi, M., Kiyama, H., Hayakawa, T., Hamada, Y. and Tsujimoto, Y. (1995) Differential expression of Notch1 and Notch 2 in developing and adult mouse brain, Mol. Brain Research, 29, 263-272.
Nakamura, S., Kawanishi, E., Nakamura, S., Watanabe, M., and Kojima, M. K. (1995) A new paralyzed flagella mutant, OC-10, in Chlamydomonas reinhardtii that can be reactivated with ATP. Phycol. Res., 43, 65-69.
Watanabe, M. (1995) Action spectroscopy : photomovement and photomorphogenesis spectra. In "CRC Handbook of Organic Photochemistry and Photobiology", (Edited by B. Horspool and P.-S. Song), CRC Press, Boca Raton, pp. 1276-1288.
II. Cooperative Research Program for the Okazaki Large Spectrograph
Erata, M., Kubota, M., Takahashi, T., Inouye, I. and Watanabe, M. (1995) Ultrastructure and phototactic action spectra of two genera of cryptophyte flagellate algae, Cryptomonas and Chroomonas. Protoplasma, 188, 258-266.
Iseki, M. and Wada, S. (1995) Action spectrum in ultraviolet region for phototropism of Bryosis rhizoids. Plant Cell Physiol. 36, 1033-1040.
Miyoshi, N., Kondo, T., Kubota, M., Ishiguro, K., Watanabe, M. and Fukuda, M. (1995) Effect of molecular formation of photosensitization for generation of active oxygen species. Photomed. Photobiol. 16, 79-82.
Nakamura, S., Kawanishi, E., Nakamura, S., Watanabe, M., and Kojima, M. K. (1995) A new paralyzed flagella mutant, OC-10, in Chlamydomonas reinhardtii that can be reactivated with ATP. Phycol. Res., 43, 65-69,
Takano, J.-I., Koizumi, H., Ohkawara, A., Kamo, N. and Ueda, T. (1995). Ultraviolet action spectra for peroxide generation in human and pig epidermail keratinocytes loaded with dihydrorhodamine 123. Arch Dermatol. Res. 287, 321-325.
Torikai, A., Chigita, K., Okisaki, F. and Nagata, M. (1995). Photo-oxidative degradation of polyethylene containing flame-retardant agents by monochromatic light. J. Appl. Polym. Sci. 58, 685-690.
Torikai, A., Hattori, A. and Eguchi, T. (1995). Wavelength effect on the photoinduced reaction of polymethylmethacrylate. J. Polymer Sci. :Part A: Polymer Chem. 33, 1867-1871.
Torikai, A., Kobatake, T., Okisaki, F. and Shuyama, H. (1995) Photodegradation of polystyrene containing flame-retardants: wavelength sensitivity and efficiency of degradation. Polymer Degradn. Stab. 50, 261-267.
Torikai, A. and Mitsuoka, T. (1995) Electron spin resonance studies of poly(methyl methacrylate) irradiated with monochromatie light. J. Appl. Polym. Sci. 55, 1703-1706.
Ueda, T. (1995). Spatiotemporal dynamics of glycolysis and cellular metabolism: toward intelligence by nonlinear chemical processes. Sensors Materials 7, 147-157.
Watanabe,M. (1995). Action spectroscopy : photomovement and photomorphogenesis spectra. In "CRC Handbook of Organic Photochemistry and Photobiology", (Edited by B. Horspool and P.-S. Song), CRC Press, Boca Raton, pp. 1276-1288.
Head: Takashi Horiuchi
Associate Professor: Kazuo Ogawa
NIBB Postdoctoral Fellow: Hiroyuki Takai
Technical Staffs: Kazuhiko Furukawa (Radiation Protection Supervisor),
Yosuke Kato (Radiation Protection Supervisor), Yoshimi Matsuda
(Radiation Protection Supervisor)
This Facility consists of a main center where a variety of radioisotopes
such as 22Na, 125I,
32P, 3H
and 14C are handled as well as
various species of Gamma-ray emitting nuclides and two subcenters,
one placed in the NIBB and the other in the NIPS. At the subcenters,
only a limited variety of radioisotopes such as 35S
and 32P are processed. The members
of the Radioisotope Facility are engaged in maintaining and controlling
both the center and subcenters, and provide users an appropriate
guidance for radioisotope handling.
In 1995, the new system for the going in and out the controlled
area was introduced. ID card has been issued to each registered
individual and non-registered persons are allowed to enter the
controlled area. In this system, when he or she goes in and out
the controlled area, his or her names, times, and places are automatically
recorded by a computer. Furthermore, the renewal of the radiation
monitoring system and the radioactive contamination assessment
system in this Facility has been supported by the secondary supplementary
budget of the Ministry of Education, Science and Culture and is
going on until the end of Mach, 1996. In these refreshed systems,
the radioactivities of air, and draining in the controlled area
of the three places will be monitored by a computer for 24 hours.
The third subcenter of this Facility will open in the next spring
(1997) in the building of the Laboratory of Gene Expression and
Regulation that is now under construction.
The teaching staffs are also engaged in their own research on
the structure and function of dynein motor protein. Dyneins are
a group of microtubule-activated ATPases that serve to convert
chemical energy into mechanical energy. They have been divided
into two large subgroups, namely, the axonemal and cytoplasmic
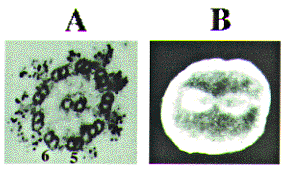
dyneins. Figure 1 shows the localization of two dyneins in the
outer arms (Ogawa et al., 1977) and the mitotic apparatus (Mohri
et al., 1976) that have been visualized by the same antibodies
directed against the motor domain of axonemal dynein (fragment
A).
The native dyneins are very large. They range in molecular mass
up to 1 to 2x106 daltons and they
are complex proteins. Each dynein contains two or three heavy
chains (HCs) with ATPase activity, which range in molecular mass
up to 500 kDa. The motor activity of dynein is associated with
these chains. Some functional differences have been reported between
HCs of outer-arm dynein. Sea urchin outer-arm dynein is a heterodimer
of HCs (Alpha and Beta) and at least the Beta-HC is able to induce
gliding of microtubules in vitro. The Alpha-HC might amplify the
function of Beta-HC and it has been reported to have no motile
activity. After the first cloning of Beta-HC from sea urchin ciliary
axonemes (Gibbons et al., 1991; Ogawa, 1991), the sequences of
HCs of axonemal and cytoplasmic dyneins from a variety of organisms
were determined in their entirety. Without exception, all the
HCs cloned to date contain four P-loop (ATP-binding) sequences
in the midregion of the molecule. Thus, they can be classified
as a four P-loop family.
The outer-arm dyneins contain two or three proteins that range
in molecular mass from 70 to 120 kDa and copurify with HCs. ICs
of sea urchin outer-arm dynein are abbreviated as IC1, IC2, and
IC3. Those of Chlamydomonas are called IC78 and IC69, and
ICs of cytoplasmic dynein are called IC74. Chlamydomonas
IC78 and IC69 were cloned by Wilkerson et al. (1995) and Mitchell
and Kang (1991),respectively. The sequences of sea urchin IC2
and IC3 were determined by Ogawa et al. (1995).

IC1 has recently been cloned (Ogawa, accession number D63884).
Rat brain IC74 was cloned by Paschal et al. (1992). Thus, all
the ICs found in the axonemal and cytoplasmic dyneins of the model
organisms used for studies of dynein function have been completely
sequenced. Comparison of amino acid sequences of IC2 and IC3 with
those of IC78 and IC69 and with that of IC74 showed that, although
all five ICs are homologous, IC2 is much more closely related
to IC78, and IC3 is much more closely related to IC69, than either
sea urchin chain or either Chlamydomonas chain is related
to each other. Regions of similarity between all five ICs are
limited to the carboxy-terminal halves of the molecules. Similarity
are due primarily to conservation of the WD repeats in all of
these chains. The WD repeats are involved in protein-protein interactions
in a large family of regulatory molecules (Neer et al., 1994).
The relative positions of WD repeats in these chains are shown
schematically in Fig. 2.
A parsimony tree for these chains (Ogawa et al., 1995) shows that,
although the carboxy-terminal halves of all of these chains contain
WD repeats, the chains can be divided into three distinct subclasses
(IC3 plus IC69, IC2 plus IC78, and IC78). By contrast, sea urchin
IC1 is not a member of the WD family. Sequence analysis showed
that IC1 consists of a thioredoxin-like (TRXL) sequence and three
nucleoside diphosphate kinase-like (NDKL) sequences. IC1 might
play a dynamic role in flagellar bending and/or wave propagation.
Ogawa, K., Kamiya, R., Wilkerson, C. G. and Witman, G. B. (1995) Interspecies conservation of outer arm dynein intermediate chain sequences defines two intermediate chain subclasses. Mol. Biol. Cell 6, 685-696.
Shimizu, H., Majima, T., Takai, H., Inaba, K. and Tomie, T. (1995) Morphological changes of wrasse sperm axoneme after their motility initiation observed with use of Atomic Force Microscopy. SPIE 2384, 45-51.
Takai, H. and Morisawa, M. (1995) Change in intracellular K concentration caused by external osmolarity change regulates sperm motility of marine and freshwater teleosts. J. Cell Sci. 108, 1175-1181.
Head of Center: Masaharu Noda
Technical Staffs: Akio Murakami, Yukiko Kabeya, Yumiko Makino,
Sonoko Ohsawa, Takeshi Mizutani
The center provides analytical instruments for biological and biophysical studies. These instruments consist of the following four groups and are supported by experienced technical staffs. Each group is equipped with instruments for general use as listed below.
Amino Acid Analyzer
HITACHI 835
Automatic Plasmid Isolation System
KURABO PI-100
DNA Sequencers
ABI 370A, 373A-36
DNA/RNA Synthesizers
ABI 381A, 392
Nucleic Acid Extractor
ABI 340A
Peptide Synthesizers
ABI 430A, 431A, 432A
Protein Sequencers
ABI 470A, 473A, 492
Capillary Electrophoresis
ABI 270A
Gas Chromatographs
SHIMADZU GC-7APTF, GC-14APFSC
Glycoprotein Analysis System
TAKARA Glyco-Tag
High Performance Liquid Chromatographs
SHIMADZU LC-10AD, 6AD
Preparative Electrophoresis System
ABI 230A
Preparative Ultracentrifuges
BECKMAN L8-80, L5-75
Table-top Ultracentrifuges
BECKMAN TL-100
Micro Preparative System
PHARMACIA SMART System
Atomic Absorption Spectrophotometer
PERKIN-ELMER 603
Dual Wavelength Spectrophotometer
HITACHI 557
EPR Spectrometer
BRUKER ER-200D
GC/Mass Spectrometer
JEOL DX-300
Inductively Coupled Plasma Spectrometer
SEIKO SPS 1200A
Infrared Spectrophotometer
JASCO A-302
Laser Raman Spectrophotometer
JASCO R-800
Light Scattering Photometer
CHROMATIX KMX-6DC
NMR Spectrometer
BRUKER AMX-360wb
Spectrofluorometers
HITACHI 850
SIMADZU RF-5000
Spectrophotometers
HITACHI 330
PERKIN ELMER Lambda Bio
Spectropolarimeter
JASCO J-40S
DELFIA Research Fluorometer
PHARMACIA
Bio Imaging Analyzers
FUJIFILM BAS 2000
Imaging Analysing Systems
KONTRON IBAS-I & II
Electrophoresis Imaging System
PDI The Discovery Series
Microscopes
CARL ZEISS Axiophot, Axiovert
Microscope Photometer
CARL ZEISS MPM 03-FL
Microdensitometer
JOYCE LOEBL 3CS