
The Department is composed of two regular divisions and two adjunct divisions. The study of this department is focused on the molecular mechanisms for the response of organisms including tissues and cells toward external and internal environments, such as light, temperature, hormones, signal-transducing substances.
Professor: Masaharu Noda
Research Associates:Nobuaki Maeda, Masahito Yamagata, Eiji
Watanabe
Institute Research Fellow:Masakazu Takahashi
Graduate Students :Hiroki Hamanaka, Junichi Yuasa, Takafumi
Shintani, Taeko Nishiwaki, Tatsunori Yamamoto*
Technical Staff :Akiko Kawai, Shigemi Ohsugi, *from
Nagoya University)
Our efforts have been devoted to studying molecular and cellular mechanisms which underlie the development of the vertebrate central nervous system. We are now seeking for the molecules and structures that regulate various cellular events in the brain morphogenesis, such as generation of neuroblasts, their migration to form the laminar structure and various nuclei, the elongation and path-finding of neural processes, and the formation and refinement of specific connections between neurons. The research has been conducted using various techniques, including molecular biology (e.g. cDNA cloning, site-directed mutagenesis), biochemistry (protein, carbohydrate), monoclonal antibodies, neuroanatomy, cell and organotypic culture, and embryo manipulation (classical embryology, gene transfer with viral vectors, and gene targeting).
Neural connection in the vertebrate brain is selective in two
ways, which we refer to as topographic and laminar. Topographic
specificity determines the orderly maps of connectivity, in which
presynaptic neurons arrayed along spatial axes project to corresponding
arrays of target cells in the target area. By contrast, laminar
specificity determines the local circuitry, in which presynaptic
neurons form synapses on the particular neurons and portions of
the neuron's surface. We have been studying the retinotectal system
in chickens to reveal the crucial factors that determine the development
of these two classes of neural connection.
Historically, most intensively studied in this system has been
the retinotopic map, where retinal ganglion cells from various
parts of the retina project topographically onto the tectal surface.
Namely, retinal axons from the temporal (posterior) retina connect
to the anterior part of the optic tectum, and nasal (anterior)
retinal axons connect to the posterior part, thereby establishing
a point-to-point projection map. Many works, since the formulation
of 'chemoaffinity theory' by Sperry, have supported the involvement
of gradient or position-specific molecules in this topographically
organized connection. Subscribing to this model, we applied a
subtractive hybridization technique, and found several novel or
identified genes to be topographically expressed along the anterior-posterior
axis in the chick retina. Among these position-specific molecules,
two molecules which belong to a same family of transcription factors
are found to be expressed in a mutually exclusive manner in either
the nasal or temporal part of the retina. Starting with these
molecules, we hope to understand the molecular mechanism by which
retinotectal topographical connection is established.
Less well studied, but striking, is a laminar selectivity of retinotectal
connection in the orthogonal direction. Retinal axons enter the
tectum through the most superficial of its 15 different laminae.
Once axons reach defined loci on the tectal surface, they branch
inwards and make connections in only 3-4 of the 15 laminae. Moreover,
each axon confines most of its synapses to one of these 'retinoreceptive'
laminae, in that neurochemically-distinguishable subsets of the
ganglion cells connect to distinct laminae to establish a functional
network. The laminar selectivity in this system may require an
axon to recognize particular target cells (cellular specificity)
and particular portions of the target cell's surface (subcellular
specificity). To identify the cellular and molecular mechanism
underlying these synaptic specificities, we are taking two interrelated
approaches. First, we are trying to establish culture systems
to analyze the recognition cues between presynaptic axons and
their target cells in vitro. The second approach
is to use molecular techniques, particularly by developing useful
probes. We have already obtained a panel of novel monoclonal antibodies
that label selectively one of 3-4 retinoreceptive layers of the
tectum during the onset of the retinotectal projection. We hope
that further characterization of these lamina-specific epitopes
will provide a clue to the mechanism that underlies the lamina-specific
synapse formation.
Developmental processes of the brain requires various cell-cell and cell-extracellular matrix interactions. In order to find key molecules in the morphogenesis of the brain, we are focusing our efforts on brain-specific proteoglycans, because proteoglycans are major extracellular matrix components in the developing brain. Previously, we identified a PBS-soluble chondroitin sulfate proteoglycan with a 300-kDa core protein, named 6B4 proteoglycan. Expression of 6B4 proteoglycan is dynamically regulated during development in various brain regions.
Recently, we cloned cDNA for 6B4 proteoglycan, and revealed that
the core protein structure corresponds precisely to the extracellular
region of a receptor-like protein tyrosine phosphatase, PTP-Zeta
(RPTPBeta). 6B4 proteoglycan is characterized by the presence
of the N-terminal carbonic anhydrase-like domain, a fibronectin
type III region, and long serine, glycine-rich domain which is
thought to be the chondroitin sulfate attachment domain. We showed
that the transmembrane form, PTP-Zeta (RPTPBeta), is also present
as a membrane-bound chondroitin sulfate proteoglycan in rat brain.
This finding suggests that these brain-specific chondroitin sulfate
proteoglycans have a role as cell signaling molecules by binding
to neural cell adhesion and extracellular matrix molecules, in
the case of 6B4 proteglycan, possibly also by competing for ligands
of the phosphatase.
In order to examine functional roles of 6B4 proteoglycan, we studied
the influence of this proteoglycan on cell adhesion and neurite
outgrowth in vitro using dissociated neurons from
the cerebral cortex and thalamus. 6B4 proteoglycan adsorbed onto
plastic tissue-culture dishes did not support neuronal cell adhesion,
but rather exerted repulsive effects on the cortical and thalamic
neurons. When neurons were densely seeded on patterned substrata
consisting of a grid-like structure of alternating poly-L-lysine
and 6B4 proteoglycan-coated poly-L-lysine domains, they were concentrated
only on the poly-L-lysine domains.
However, 6B4 proteoglycan did not retard the differentiation of
neurons but rather promoted neurite outgrowth and development
of the dendrites of cortical neurons, when neurons were sparsely
seeded on poly-L-lysine-conditioned coverslips continuously coated
with 6B4 proteoglycan (Fig 1.).
This effect of 6B4 proteoglycan on the neurite outgrowth of cortical
neurons was apparent even on coverslips co-coated with fibronectin
or tenascin. By contrast, the neurite extension of thalamic neurons
was not modified by 6B4 proteoglycan. Chondroitinase ABC or keratanase
digestion of 6B4 proteoglycan did not affect its neurite outgrowth
promoting activity, suggesting that a protein moiety is responsible
for the activity. 6B4 proteoglycan transiently promoted tyrosine
phosphorylation of an 85-kDa protein in the cortical neurons but
not in the thalamic neurons, which correlated with the induction
of neurite outgrowth. These results suggest that 6B4 proteoglycan
modulates morphogenesis and differentiation of neurons spatiotemporally
depending on cell types.
Drescher, U., Kremoser, C. Handwerker, C., Löschinger, J., Noda, M. and Bonhoeffer, F. (1995) In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for Eph receptor tyrosine kinases. Cell 82, 359-370.
Leber, S. M., Yamagata, M. and Sanes, J. R. (1996) Gene transfer using replication-defective retroviral and adenoviral vectors. In Methods in Avian Embryology (Bronner-Fraser, M. ed., Methods in Cell Biology, Academic Press, Orlando) 51, pp. 161-183.
Maeda, N. and Noda, M. (1996) 6B4 proteoglycan/phosphacan is a repulsive substratum but promotes morphological differentiation of cortical neurons. Development 122, 647-658.
Maeda, N., Hamanaka, H., Oohira, A. and Noda, M. (1995) Purification, characterization and developmental expression of a brain-specific chondroitin sulfate proteoglycan, 6B4 proteoglycan/phosphacan. Neuroscience 67, 23-35.
Nishizuka, M., Ikeda, S., Arai, Y., Maeda, N. and Noda, M. (1996) Cell surface-associated extracellular distribution of a neural proteoglycan, 6B4 proteoglycan/phosphacan, in the olfactory epithelium, olfactory nerve, and cells migrating along the olfactory nerve in chick embryos. Neurosci. Res., in press.
Watanabe, E., Maeda, N., Matsui, F., Kushima, Y., Noda, M. and Oohira, A. (1995) Neuroglycan C, a novel membrane-spanning chondroitin sulfate proteoglycan that is restricted to the brain. J. Biol. Chem. 270, 26876-26882.
Watanabe, E., Matsui, F., Keino, H., Ono, K., Kushima, Y., Noda, M. and Oohira, A. (1996) A membrane-bound heparan sulfate proteoglycan that is transiently expressed on growing axons in the rat brain. J. Neurosci. Res., in press.
Yamagata, M. and Sanes, J. R. (1995) Target-independent diversification and target specific projections of chemically coded retinal ganglion cell. Development 121, 3763-3776.
Yamagata, M., Herman, J. -P. and Sanes, J. R. (1995) Lamina-specific expression of adhesion molecules in developing chick optic tectum. J. Neurosci. 15, 4556-4571.
Professor: Norio Murata
Associate Professor:Hidetoshi Iida
Research Associates:Ikuo Nishida, Dmitry A. Los, Atsushi Sakamoto,
Yoshitaka Nishiyama
Monbusho Foreign Scientists: Laszlo Mustardy1),
Malay Kumar Ray2), Alia3)
JSPS Postdoctoral Fellow: Michael P. Malakhov4)
NIBB Fellow:Iwane Suzuki
JSPS Visiting Scientists: Rajinder S. Dhindsa5),
Dilley Richard 6), Ding-Ji Shi7),
Éva Hideg1),
P. Pardha Saradhi3), Nelly Tsvetkova8),
Diego de Mendoza9)
Visiting Scientists: Yasushi Tasaka, Makiko S. Okumura, Zoltan
Gombos1), George Papageorgiou10),
Prasanna Mohanty11), Ferenc Lajko1),
Julian Eaton-Rye12), Ashraf S.
Haider13), Estelle Delphin14)
Graduate Students:Katsuzo Noguchi, Patcharaporn Deshnium, Sayamrat
Panpoom, Masami Inaba, Hiroshi Yamamoto
Technical Staffs:Sho-Ichi Higashi, Miki Ida
(1) from Biological Research
Center, Szeged, Hungary)
(2) from Centre for Cellular
and Molecular Biology, Hyderabad, India)
(3) from Jamia Millia
Islamia University, New Delhi, India)
(4) from Plant Physiology
Institute, Moscow, Russia)
(5) from McGill University,
Montreal, Canada)
(6) from Purdue University,
Lafayette, IN, U.S.A.)
(7) from Institute of
Botany, Academia Sinica, Beijing, China)
(8) from
Bulgarian Academy of Sciences, Sofia, Bulgaria)
(9) from Universidad
Nacional de Rosario, Rosario, Argentina)
(10) from National Center
for Scientific Research Demokritos, Athens, Greece)
(11) from Jawaharlal
Nehru University, New Delhi, India)
(12) from Otago University,
Dunedin, New Zealand)
(13) from Agricultural
Genetic Engineering Research Institute, Giza, Egypt)
(14) from Laboratoire
de Photorégulation
et Dynamique des Membranes Végétales,
Paris, France)
The research effort of this division is directed toward a full understanding of the molecular mechanisms that allow plants to acclimate to and tolerate extremes of temperature and salinity. In 1995, significant progress was made in the following areas as a result of studies with cyanobacteria and higher plants as experimental materials.
Living organisms are exposed to changes in ambient temperatures.
However, they are able to maintain the appropriate fluidity of
their membrane lipids by regulating the extent of unsaturation
of their fatty acids. For example, cyanobacterial cells respond
to a decrease in temperature by introducing double bonds into
the fatty acids of membrane lipids, thereby compensating for the
temperature-induced decrease in the molecular motion of membrane
lipids. Desaturases catalyze the introduction of these specific
double bonds. We have demonstrated that the low temperature-induced
desaturation of fatty acids in membrane lipids is regulated at
the level of expression of the genes for the desaturases.
We have shown that the levels of the transcripts of the desA,
desB, and desD genes (for the Delta12, Delta15 and
Delta6 desaturases, respectively) increase about 10-fold after
shift in culture temperature from 34°C
to 22°C. However, the
level of the transcript of the desC gene (for the Delta9
desaturase) remains constant after such a temperature shift, suggesting
that expression of this gene is constitutive. Sequencing of the
nucleotides at the 5' termini of the genes for all four desaturases
indicated that the sites of initiation of transcription are quite
similar among all four genes. Identification of the regulatory
elements of these genes and cloning of the relevant trans-acting
factors are in progress.
As part of our efforts to understand the role of unsaturation of membrane lipids, we disrupted the desA and desD genes in Synechocystis sp. PCC 6803 by inserting antibiotic-resistance gene cartridges. This disruption greatly reduced the extent of unsaturation of the fatty acids of the membrane lipids. In the wild-type strain, the saturated, mono-unsaturated, di-unsaturated, and tri-unsaturated lipid molecules each accounted for 15% of the total membrane lipids. By contrast, in the desD- mutant, the triunsaturated lipid molecules were absent, while in the desA-/desD- mutant, 80% of the total membrane lipids were monounsaturated and only 15% were fully saturated. These decreases in the unsaturation of membrane lipids greatly reduced the tolerance of the cyanobacterium to low temperature in the light. These results, together with other observations, suggest that unsaturated fatty acids play an important role in protection of the photosynthetic machinery against low-temperature photoinhibition. The extent of photoinhibition in intact cells reflects a balance between the light-induced inactivation of the photosystem II protein complex and the recovery of the complex from the inactivation. Therefore, we were able to separate the two processes using inhibitors of protein synthesis. Our results demonstrated that the unsaturation of membrane lipids had no effect on the light-induced inactivation process but did accelerate the recovery of the photosystem II complex from the photoinactivated state. Further analysis indicated that the unsaturation of membrane lipids accelerated the incorporation of the precursor to the D1 protein (an important protein component of the photosystem II complex) or the processing of the precursor to the D1 protein to generate the mature D1 protein.
We also focused on the responses of plants to high-temperature
stress. Photosynthesis is one of the physiological processes in
plants that is most susceptible to heat stress. Thus, stabilization
of the photosynthetic machinery against heat helps plants to tolerate
higher temperatures.
We have studied the molecular mechanism responsible for the stabilization
against heat of oxygen evolution in the cyanobacterium Synechococcus
sp. PCC 7002. When thylakoid membranes isolated from the cyanobacterial
cells were treated with a low concentration of Triton X-100, the
stability to heating of the evolution of oxygen decreased markedly.
From the extracts obtained by this treatment with Triton X-100,
we purified two proteins that were able to restore the heat stability
of oxygen evolution. One protein was identified as cytochrome
c-550, having a low redox potential. The other protein
of 13 kDa was identified as a homology of the 9-kDa extrinsic
protein of cyanobacterial photosystem II. These results indicate
that cytochrome c-550 and the 13-kDa protein are involved
in the stabilization against heat of the evolution of oxygen and,
therefore, in the stability of the photosynthetic machinery at
elevated temperatures. We isolated the genes for cytochrome c-550
and the 13-kDa protein of Synechococcus sp. PCC 7002. Disruption
of these genes in Synechococcus sp. PCC 7002 is now in
progress in our laboratory. We have also studied the effects of
elevated temperatures on photosynthesis in cultured soybean cells.
The heat stability of oxygen evolution in these cells was enhanced
by 3°C upon an increase
in the growth temperature from 25°C
to 35°C.
When cultured under saline conditions, some plants produce compatible solutes to avoid the deleterious effects of external salt. Glycinebetaine, a quaternary ammonium compound, is one such solute that is found in halotolerant plants and bacteria. To examine the effect of glycinebetaine on photo-synthesis in vivo, we transformed the salinity-sensitive cyanobacterium Synechococcus sp. PCC 7942 with the codA gene for choline oxidase from Arthrobacter globiformis, which catalyzes the oxidation of choline to glycinebetaine. The transformed cells accumulated glycinebetaine at intracellular levels of 60-80 mM and simultaneously acquired the ability to tolerate salt stress. We also observed that the transformed cells could grow at 20°C, a temperature that markedly retarded the growth of the wild-type cells. Photosynthesis in the transformed cells was more resistant to photoinhibition at low temperatures than that in wild-type cells. Furthermore, the recovery of photosystem II from photoinhibition at low temperatures was enhanced in the transformed cells. These findings suggest that glycinebetaine enhanced the growth of the transformed cells at 20°C by accelerating the recovery of the photosynthetic machinery from photoinhibition.
A higher plant, Arabidopsis thaliana, was transformed with the codA gene for choline oxidase under control of the 35S promoter of cauliflower mosaic virus. Such transformation allowed the plant to accumulate glycinebetaine and enhanced its ability to tolerate salt and cold stresses. When cultured in the presence of 100 mM NaCl, transformed plants grew well whereas wild-type plants failed to grow. The transformed plants tolerated 200 mM NaCl, which was lethal to wild-type plants. After plants had been incubated with 400 mM NaCl for two days, the photosystem II activity of wild-type plants had almost completely disappeared, whereas that of transformed plants remained at more than 50% of the original level. When exposed to low temperatures in the light, leaves of wild-type plants exhibited symptoms of chlorosis, whereas those of transformed plants did not. Thus, the genetic modification of Arabidopsis thaliana that allowed it to accumulate glycinebetaine also enhanced its ability to tolerate both salt and cold stresses.
Avelange-Macherel, M. H., Macherel, D., Wada, H. and Murata, N. (1995) Site-directed mutagenesis of histidine in the Delta12 acyl-lipid desaturase of Synechocystis. FEBS Lett. 361, 111-114.
Deshnium, P., Los, D. A., Hayashi, H., Mustardy, L. and Murata, N. (1995) Transformation of Synechococcus with a gene for choline oxidase enhances tolerance to salt stress. Plant Mol. Biol. 29, 897-907.
Fukuchi-Mizutani, M., Savin, K., Cornish, E., Tanaka, Y., Ashikari, T., Kusumi, T. and Murata, N. (1995) Senescence-induced expression of a homologue of Delta9 desaturase in rose petals. Plant Mol. Biol. 29, 627-635.
Iida, H., Ohya, Y. and Anraku, Y. (1995) Calmodulin-dependent protein kinase II and calmodulin are required for induced thermo-tolerance in Saccharomyces cerevisiae. Curr. Genet. 27, 190-193.
Ishizaki-Nishizawa, O., Azuma, M., Ohtani, T., Murata, N. and Toguri, T. (1995) Nucleotide sequence of cDNA from Spinacia oleracea encoding plastid glycerol-3-phosphate acyl-transferase. Plant Physiol. 108, 1342.
Kanervo, E., Aro, E.-M. and Murata, N. (1995) Low unsaturation level of thylakoid membrane lipids limits turnover of the D1 protein of photosystem II at high irradiance. FEBS Lett. 364, 239-242.
Malakhov, M. P., Los, D. A., Wada, H., Semenenko, V. E. and Murata, N. (1995) Characterization of the murF gene of the cyanobaterium Synechocystis sp. PCC 6803. Microbiology 141, 163-169.
Moon, B. Y., Higashi, S., Gombos, Z. and Murata, N. (1995) Unsaturation of chloroplastic membrane-lipids protects photosynthesis against low-temperature photoinhibition in transgenic tobacco plants. Proc. Natl. Acad. Sci. U.S.A. 92, 6219-6223.
Tanida, I., Hasegawa, A., Iida, H., Ohya, Y. and Anraku, Y. (1995) Cooperation of calcineurin and vacuolar H+-ATPase in intracellular Ca2+ homeostasis of yeast cells. J. Biol. Chem. 270, 10113-10119.
Murata N. and Wada H. (1995) Acyl-lipid desaturases and their importance in the tolerance and acclimatization to cold of cyanobacteria. Biochem. J. 308, 1-8.
Papageorgiou G. C. and Murata N. (1995) The unusually strong stabilizing effect of glycinebetaine on the structure and function of the oxygen-evolving photosystem II complex. Photosyn. Res. 44, 243-252.
Deshnium, P., Los, D. A., Hayashi, H. and Murata, N. (1995) Glycinebetaine enhances tolerance to salt stress in transgenic cyanobacterium. In Photosynthesis: From Light to Biosphere (P. Mathis, ed.), Kluwer Academic Publishers, Dordrecht, Vol. IV, pp. 637-640.
Murata, N. (1995) Genetic engineering of photosynthetic capability under temperature and salinity stresses. In Photosynthesis: From Light to Biosphere (P. Mathis, ed.), Kluwer Academic Publishers, Dordrecht, Vol. IV, pp. 763-770.
Papageorgiou, G. C. and Murata, N. (1995) Thylakoids from salt-stressed Spinacea oleracea are more active in photosynthetic oxygen evolution and more tolerant to alkaline pH than thylakoids from unstressed plants. In Photosynthesis: From Light to Biosphere (P. Mathis, ed.), Kluwer Academic Publishers, Dordrecht, Vol. IV, pp. 645-648.
Professor (adjunct): Kimiyuki Satoh
Associate Professor (adjunct): Hirokazu Kobayashi
Research Associate: Noritosi Inagaki
Graduate Students: Kyoichi Isono*, Yoshihiro Narusaka*,
Tatsuya Tomo*, Satoshi Matsumoto*, Akihiko
Tohri*, (*from Okayama University)
From an energetic point of view, the conversion of light energy
into chemical energy in photosynthesis is the most important biological
process on earth. The highly efficient energy conversion in this
process is ensured by the highly ordered organization of molecules
in the photochemical reaction center, in a physical, chemical
and biological sense. The project in this division is aiming to
elucidate the organization of photosystem II reaction center of
oxygenic photosynthesis which has a unique property to generate
a strong oxidant for utilizing the water molecule as electron
donor.
In the first approach, molecular organization of the photosystem
II reaction center, which has been identified in our study, will
be analyzed by several methods which include crystallographic
analysis, chemical modification & cross-linking analysis and
optical & EPR spectroscopies. Structure-functional analysis
will also be conducted for the reaction center using random and
site-directed mutagenesis for transformable algae, Synechocystis
PCC 6803 and Chlamydomonas reinhardtti. The principal target
of these analyses will be the structure and molecular environment
of P-680, the primary donor, which determine the redox potential
of this unique photosystem.
In the second approach, the effort will be focused on the elucidation
of molecular mechanism of light-regulated metabolic turnover of
a subunit of the photosystem II reaction center, D1 protein. Some
of unique steps are involved in this process; i.e., photo-damage
of the photochemical reaction center, specific degradation of
the impaired protein subunit, light-regulated gene expression
at the translational level, post-translational cleavage of the
C-terminal extension and the incorporation of cofactors and subunits
into a multi-component pigment-protein complex.
(1) A photochemically inactive pheophytin (Pheo) a molecule in the photosystem II reaction center, which contains 6 chlorophyll a and two Beta-carotene molecules for every two Pheo a, was identified by fluorescence excitation spectroscopy at 77K, and confirmed by absorption, linear dichroism, and magnetic circular dichroism spectra (in collaboration with Dr. M. Mimuro, Div. of Bioenergetics). When the emission was monitored at 740 nm, photochemically active Pheo a was detected as peaks at 418, 513, 543, and 681 nm. However, when the emission was monitored at 665 nm, two bands were observed at 414 and 537 nm; these bands were insensitive to photochemical reduction of the primary acceptor and thus appear to be due to photochemically inactive Pheo a.
(2) Solvent extraction at�`4°C in complete darkness, and subsequent analysis by high-pressure liquid chromatography using an apparatus equipped with a two-dimensional diode-array detector, spectroscopically identified 15-cis-Beta-carotene in the photosystem II reaction center. The result revises a previous conclusion that Beta-carotene bound to the reaction center takes the all-trans configuration, and generalizes the concept established for purple photosynthetic bacteria that 15-cis-carotenoid is naturally selected by the photosynthetic reaction centers (in collaboration with Drs. Y, Koyama et al., Kwansei Gakuin University).
(3) Site-directed modifications of the environment of the primary
donor of photosystem II reaction center were attempted, using
Synechocystis PCC 6803, in order to elucidate molecular
interactions responsible for the high redox-potential.
(1) The D1 subunit of the photosystem II reaction center has a
C-terminal extension consisting of 8-16 amino acids. Post-translational
removal of this part of the protein is essential for the manifestation
of oxygen-evolving function in photosystem II. The enzyme involved
in the C-terminal processing of the D1 precursor protein (pD1)
of the photosystem II reaction center was purified from extracts
of sonicated spinach thylakoids by a method that included chromatography
on quaternary aminoethyl anion-exchange, hydroxyl-apatite, copper-chelating
affinity and gel-filtration columns. The enzyme was identified,
from its chromatographic behavior, to be a monomeric protein of
about 45 kDa. The sequence of the amino-terminal 27 amino acids
of this protein was determined directly, which exhibited low but
appreciable (37%) homology to that deduced from a gene (ctpA)
in Synechocystis PCC 6803 that was proposed recently to
encode the processing protease form results of genetic complementation
analysis.
Based on the amino acid sequence data of the purified protease,
a cDNA clone encoding the enzyme has identified and sequenced,
from a spinach green leaf cDNA library. By the analysis, the full-length
transcript was established to consist of 1906 nucleotides and
a poly(A) tail, containing an open reading frame corresponding
to a protein with 539 amino acid residues. By comparing the amino
acid sequence of the purified protease with that deduced form
nucleotide sequence of the cDNA clones, the enzyme was shown to
be furnished with an extra N-terminal extension characteristic
of both a transit peptide and a signal sequence. This suggests
that the protease is synthesized in the cytosol and translocated
into the lumenal space of thylakoids. The mature part of the enzyme
consists of 389 amino acid residues and exhibits a significant
sequence homology with two groups of proteins as demonstrated
by a computer homology search; i.e., (i) the deduced sequence
of a protein proposed to be the C-terminal processing protease
for pD1 in Synechocystis PCC 6803 and (ii) proteases for
C-terminal cleavage identified in Escherichia coli and
Bartonella bacilliformis.
(2) The synthesis of D1 precursor protein is regulated by light at the stage of translation. The mechanism of this light regulation was analyzed using isolated pea chloroplasts. Accumulation of the precursor and the mature form of the D1 protein of the photosystem II reaction center in illuminated pea chloroplasts was prevented by the addition of the inhibitors of photosynthetic electron transport, i.e., atrazine, 3-(3,4-dichlorophenyl-1,1-dimethylurea and bromoxynil. Under such conditions, the compensatory accumulation of two translational intermediates of the D1 protein of 22 and 24 kDa was induced by the addition of ATP, as also observed in darkness in the presence of ATP (Taniguchi et al., FEBS Lett. 317, 57-61, 1993), suggesting that the synthesis of the full-length D1 protein requires a factor that is generated by the operation of photosynthetic electron transport. The accumulation of the full-length D1 protein was induced in the light, even in the presence of atrazine, when both 2,6-dichlorophenolindophenol and ascorbate were also present and in darkness upon the addition of dithiothreitol. Moreover, reagents with a relatively low redox potential, namely, duroquinone and methylviologen, prevented the accumulation. These observations suggest that the translation of the D1 protein is regulated at specific steps during the elongation of the polypeptide via a redox change in a component around photosystem I.
(3) Photo-tolerant mutants of Synechocystis PCC 6803 were obtained by in vitro random mutagenesis of psbAII (gene for D1 protein) by PCR under a condition for reduced fidelity of amplification. A preliminary characterization has been carried out for a mutant. The mutant strain (G-3) was shown to have five substitutions in the amino acid in regions other than so-called "PEST-like sequence" in the D1 protein, i.e., Phe-260 and Phe-273 to Ser, Gln-304 and Gln-310 to Arg, and Bal-351 to Ala. The mutant cells were tolerant to high irradiance both in terms of pigmentation and viability(Fig. 1). The functional characterization of photosynthesis in the mutant has been conducted.

Bialek-Bylka, G.E., Tomo, T., Satoh, K. and Koyama, Y. (1995) 15-cis-Beta-Carotene found in the reaction center of spinach photosystem II. FEBS Lett. 363, 137-140.
Fujita, S., Inagaki, N., Yamamoto, Y., Taguchi, F., Matsumoto, A. and Satoh, K. (1995) Identification of the carboxyl-terminal processing protease for D1 precursor protein of photosystem II reaction center of spinach. Plant Cell Physiol. 36, 1169-1177.
Mimuro, M., Tomo, T., Nishimura, Y., Yamazaki, I. and Satoh, K. (1995) Identification of a photochemically inactive pheophytin molecule in spinach D1-D2-cyt b559 complex. Biochem. Biophys. Acta, in press.
Nozawa, T., Kobayashi, M., Wang, Z.-Y., Itoh, S., Iwaki, M., Mimuro, M. and Satoh, K. (1995) Magnetic circular dichroism investigation on chromophores in reaction centers of photosystem I and II of green plant photosynthesis. Spectrochimica Acta 51A, 125-134.
Satoh, K. (1995) Introduction to photosystem II reaction center -Isolation and some biochemical and biophysical characterization-. in "Oxygenic Photosynthesis: The Light Reactions" (D.R. Ort & C.F. Yocum eds.), Kluwer Academic Publishers (Dordrecht), in press.
Taguchi, F., Yamamoto, Y. and Satoh, K. (1995) Recognition of the structure around the site of cleavage by the carboxyl-terminal processing protease for D1 precursor protein of the photosystem II reaction center. J. Biol. Chem. 270, 10711-10716.
Professor: Masatoshi Takeichi
Research Associates: Akinao Nose, Kazuaki Tatei
Institute Research Fellow: Emiko Shishido
Post-doctoral Fellow: Takako Isshiki
Graduate Students: Tatsuo Umeda, Hiroki Taniguchi, Takeshi
Umemiya (from Kyoto University)
How individual nerve cells find and recognize their targets during
development is one of the central issues in modern biology. The
aim of our division is to elucidate the molecular mechanism of
axon guidance and target recognition by using the simple and highly
accessible neuromuscular system of Drosophila.
The musculature of Drosophila embryos consists of 30 identifiable
muscle fibers per hemisegment. Each muscle fiber is innervated
by a few motoneurons in a highly stereotypic manner. The high
degree of precision and previous cellular manipulations of neuromuscular
connectivity suggest the presence of recognition molecules on
the surface of specific muscle fibers which guide the growth cones
of motoneurons. We have previously isolated several enhancer trap
lines that express the reporter gene Beta-galactosidase (Beta-gal)
in small subsets of muscle fibers prior to innervation. By molecularly
characterizing these lines, we are trying to identify genes that
play roles in the specification of the muscles and neuromuscular
connectivity. Previous studies showed that two of the lines are
insertions in the connectin and Toll genes, that
encode cell recognition molecules which belong to the leucine-rich
repeat (LRR) family. We are currently studying the function of
these genes, and also characterizing other lines by molecular
and genetic analysis.
Connectin is expressed on a subset of muscle fibers (primarily
lateral muscles) and on the axons, growth cones of the motoneurons
which innervate these muscles (primarily SNa motoneurons) and
on several associated glial cells. When coupled with its ability
to mediate homophilic cell adhesion in vitro, these results led
to the suggestion that Connectin functions as an attractive signal
for SNa pathfinding and targeting.
To study the role of Connectin in vivo, we ectopically expressed
Connectin on all muscles by using MHC (myosin heavy chain) promoter
(MHC-connectin) in the P-element mediated transformants.
In MHC-connectin, SNa nerves were observed to send extra
axon branches that form ectopic nerve endings on muscles 12, muscles
they would never innervate in wild type. This phenoype was highly
penetrant and was observed in over 60% of the segments examined.
Furthermore, the ectopic innervation on muscle 12 was dependent
on the Connectin expression on SNa. These results showed that
Connectin functions as an attractive and homophilic guidance molecule
for SNa in vivo.
We are conducting molecular and genetic analysis of other enhancer
trap lines that are expressed in small subsets of muscles and/or
neurons. One of them, P750 expresses Beta-gal in subsets of ventral
and dorsal muscles and in a small number of neurons in the central
nervous system (CNS). The cDNA cloning and sequencing revealed
that P750 encodes a novel transmembrane protein that belongs to
LRR family. It is interesting that three of the five muscle enhancer
trap lines that we have thus far characterized contain LRRs. It
may point to the important role of this motif in the development
of muscles and/or neuromuscular specificity. Within the LRR family,
P750 was found to be most related to the Drosophila tartan
gene that have been implicated in neural and muscular development.
To study the function of P750, we have isolated the loss-of-function
mutants, and are currently characterizing the phenotype.
Another enhancer trap line, AN34 expresses Beta-gal in a single
muscle fiber (#18) per hemisegment (Fig.1A). The remarkable specificity
in its expression pattern (one out of 30 muscle fibers) makes
it a good candidate for the muscle target recognition molecule.
AN34 was found to encode a secreted protein with extensive amino
acid similarity to rat F-spondin. F-spondin is a secreted molecule
expressed at high levels in the floor plate and has been shown
to promote neural cell adhesion and neurite extension in vitro.
These results strongly suggest that AN34 is involed in motoneuronal
gidance and/or targeting.
There are three regions that are highly conserved between AN34
and F-spondin. One of them (domain III) is a known repeating motif
called thrombospondin type I repeats (TSRs). The other two (domain
I and II) are novel conserved sequences that we identified. The
amino acid identity in domain II is particularly high suggesting
its functional importance. We thus searched for novel genes in
Drosophila that contain this domain by using PCR, and succeeded
in cloning two new genes. Similarly, Higashijima et al. (Div.
of Cellular Communication) cloned two novel genes in addition
to F-spondin in the zebrafish. All of these newly cloned genes
share similar overall structure with AN34 and F-spondin in that
they possessed domain I, II and one to six TSRs.
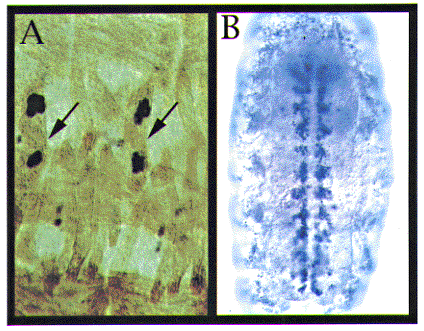
(A) AN34 enhancer trap line express Beta-galactosidase (Beta-gal) in the nuclei of single muscle fiber #18 (arrows) in each hemisegment. Anti Beta-gal antibody staining in blue and general muscle staining (as visualized with a monoclonal antibody 27D) in brown.
(B) #8 in situ hybridization. #8 mRNA is expressed in a
small subset of glia along the longitudinal connectives.
They thus constitute a novel gene family of secreted protein with
potential roles in neural adhesion.
One of the newly identified genes in Drosophila (tentatively
called #3) is expressed in a subset of hemocytes, secretory cells
of the extracellular matrix (ECM) proteins. Another (called #8)
is expressed in a subset of glia that sit along the longitudinal
axon tracts in the CNS (Fig.1B).
The specificity of their expression suggest that they might be
involved in the guidance of specific axons. We are currently trying
to isolate the loss-of-function mutants of these three genes (AN34,
#3, #8) as well as the transgenic flies that ectopically express
these molecules (as described for Connectin) to study their roles
in neural development.
Another line rH96 is expressed in the precursors of dorsal muscles. It is also expressed in a subset of neuroblasts and glioblasts in the CNS. In particular, it is expressed in the longitudinal glioblasts (LGB), precursor cells for the longitudinal glia whose lineage is well characterized. rH96 was shown to encode a putative transcription factor with a homeobox. Analysis of the loss-of-function mutants revealed severe defects in the formation of muscles and the CNS. The dorsal muscles that normally express the molecule were often missing totally or failed to form properly. The overall structure of the CNS axon tract was also seriously disrupted. Close examination of the development of the LGB leneage using specific markers showed that the glioblasts fail to divide and migrate properly in the mutants. These results suggested that rH96 play essential roles in the specification and/or differentiation of precursors for neurons, glia and muscles. We are currently trying to ectopically express rH96 to characterize more in detail the function of rH96 in the development of muscles and the nervous system.
Kimura, Y., Matsunami, H., Inoue, T., Shimamura, K., Uchida, N., Ueno, T., Miyazaki, T. and Takeichi, M. (1995) Cadherin-11 expressed in association with mesenchymal morphogenesis in the head, somite, and limb bud of early mouse embryos. Develop. Biol. 169, 347-358.
Matsunami, H. and Takeichi, M. (1995) Fetal brain subdivisions defined by R- and E-cadherin expressions: evidence for the role of cadherin activity in region-specific, cell-cell adhesion. Develop. Biol. 172, 466-478.
Nakagawa, S. and Takeichi, M. (1995) Neural crest cell-cell adhesion controlled by sequential and subpopulation-specific expression of novel cadherins. Development 121, 1321-1332.
Overduin, M., Harvey, T.S., Bagby, S., Tong, K.I., Yau, P., Takeichi, M. and Ikura, M. (1995) Three-dimensional solution structure and calcium interaction of the epithelial cadherin domain responsible for selective adhesion. Science 267, 386-389.
Shibamoto, S., Hayakawa, M., Takeuchi, K., Hori, T., Miyazawa, K., Kitamura, N., Johnson, K.R., Wheelock, M.J., Matsuyoshi, N., Takeichi, M. and Ito, F. (1995) Association of p120, a tyrosine kinase substrate, with E-cadherin/catenin complexes. J. Cell Biol. 128, 949-957.
Shibuya, Y., Mizoguchi, A., Takeichi, M., Shimada, K. and Ide, C. (1995) Localization of N-cadherin in the normal and regenerating nerve fibers of the chicken peripheral nervous system. Neuroscience 67, 253-261.
Shiozaki, H., Kadowaki, T., Doki, Y., Inoue, M., Tamura, S., Oka, H., Iwazawa, T., Matsui, S., Shimaya, K., Takeichi, M. and Mori, T. (1995) Effect of epidermal growth factor on cadherin-mediated adhesion in a human oesophageal cancer cell line. British J. Cancer 71, 250-258.
Takeichi, M. (1995) Morphogenetic roles of classic cadherins. Current Opinion Cell Biol. 7, 619-627.