
Professor:Mikio Nishimura
Research Associates: Makoto Hayashi, Ikuko Hara-Nishimura,
Tomoo Shimada
NIBB Postdoctral Fellow: Akira Kato
Graduate Students: Tetsu Kinoshita, Nagako Hiraiwa, Shouji
Mano, Masahiro Aoki 1), Daigo Takemoto 2)
Technical Staffs: Maki Kondo, Katsushi Yamaguchi, Yasuko Koumoto
Visiting Scientists: Kyoko Hatano 3)
1) from Shinshu University
2) from Nagoya University
3) from Kyoto Univerisity
Higher plant cells contain several distinct organelles that play
vital roles in cellular physiology. During proliferation and differentiation
of the cells, the organelles often undergo dynamic changes. The
biogenesis of new organelles may occur, existing organelles may
undergo a transformation of function, and other organelles may
degenerate. Because the dynamic transformation of organellar function
(differentiation of organelles) is responsible for the flexibility
of differentiation events in higher plant cells, the elucidation
of regulatory mechanisms underlying organelle transformation are
currently being studied in this division.
Dramatic metabolic changes which underlie the shift from heterotrophic to autotrophic growth occur during the in greening of seeds Accompanying these metabolic changes, many constitutive organelles are functionally transformed. For example, etioplasts differentiate into chloroplasts and mitochondria acquire the ability to oxidize glycine. Glyoxysomes, which are microbodies engaged in the degradation of reserve oil via b-oxidation and the glyoxylate cycle, are transformed into leaf peroxisomes that function in several crucial steps of photorespiration. After the functional transition of glyoxysomes to leaf peroxisomes during the greening of pumpkin cotyledons, the reverse microbody transition of leaf peroxisomes to glyoxysomes occurs during senescence. To clarify the molecular mechanisms underlying the microbody transition, the change with development in microbody membrane proteins during transformation of glyoxysomal to leaf peroxisomes was characterized. Two proteins in glyoxysome membranes, with molecular masses of 31 and 28 kDa, were purified and characterized. The 31-kDa membrane protein was found to be a novel isoenzyme of ascorbate peroxidase. Intact glyoxysomes and leaf peroxisomes had no latent peroxidase activity, an indication that the active site of the ascorbate peroxidase was exposed to the cytosol and that the peroxidase would scavenge hydrogen peroxide leaked from microbodies (Fig. 1).
Fig. 1. Possible function of the ascorbate peroxidase localized on microbody membranes. mbAPX, microbody ascorbate peroxidase; cAPX, cytosolic ascorbate peroxidase; CAT, catalase; AsA, ascorbate; DHA, dehydroascorbate; MDA, monodehydroascorbate.
Analysis of these membrane proteins during development revealed that the amounts of these membrane proteins decreased during the microbody transition and that the larger one was retained in leaf peroxisomes, whereas the smaller one could not be found in leaf peroxisomes after completion of the microbody transition. These results clearly showed that membrane proteins in glyoxysomes change dramatically during the microbody transition, as do the enzymes in the matrix.
Citrate synthase catalyzes the conversion of oxaloacetate to citrate. In plant cells, there are two isozymes of citrate synthase, one is an enzyme involved in the glyoxylate cycle in glyoxysomes (gCS) and the other is an enzyme involved in the TCA cycle in mitochondria. To study the structural characteristics and development of gCS during the microbody transition at the molecular level, we cloned cDNA for pumpkin gCS. The deduced amino acid sequence of gCS did not have a typical peroxisomal targeting signal at its carboxyl terminus. A study of the in vitro expression of the cDNA and an analysis of the amino-terminal sequence of the citrate synthase indicated that gCS is synthesized as a larger precursor that has a cleavable amino-terminal presequence of 43 amino acids. The predicted amino-terminal sequence of pumpkin gCS was highly homologous to those of other microbody enzymes, such as 3-ketoacyl-CoA thiolase of rat and malate dehydrogenase of watermelon that are also synthesized as precursors of higher molecular mass. An immunoblot analysis showed that the level of gCS protein increased markedly during germination and decreased rapidly during the light-induced transition of microbodies from glyoxysomes to leaf peroxisomes. By contrast, the level of mRNA for gCS reached a maximum earlier than that of the protein and declined even in darkness. The level of the mRNA was low during the microbody transition. These results indicate that the accumulation of the gCS protein does not correspond to that of the mRNA and that degradation of gCS is induced during the microbody transition, suggesting that post-transcriptional regulation plays an important role in the microbody transition.
During seed maturation, protein-storage vacuoles are converted
to protein bodies that are found in dry seeds. In contrast, during
the postgermination growth of seeds, protein bodies fuse with
one another and are converted to a central vacuole. To investigate
this transition, we prepared protein-body membranes from dry seeds
of pumpkin (Cucurbita sp.) and characterized their protein
components. Five major proteins (designated MP23, MP27, MP28,
MP32 and MP73) were detected in the protein-body membranes. We
have isolated cDNAs for MP23, MP27, MP28 and MP32 and characterized
them.
Among the five membrane proteins, MP27 and MP32 disappeared most
rapidly during seedling growth. Both MP27 and MP32 were encoded
by a single cDNA. The deduced precursor polypeptide was composed
of a hydrophobic signal sequence, MP27 and MP32, in that order.
A putative site of cleavage between MP27 and MP32 was located
on the COOH-terminal side of asparagine 278, an indication that
the post-translational cleavage may occur by the action of a vacuolar
processing enzyme (VPE) that converts proprotein precursors of
seed proteins into the mature forms. Immunoelectron microscopic
analysis showed that MP27 and MP32 was associated with the protein-body
membrane of dry pumpkin seeds. The degradation of MP27 and MP32
starts just after seed germination and proceeds in parallel with
the transformation of the protein bodies into vacuoles.
Molecular characterization revealed that both MP28 and MP23 belong
to the seed TIP (tonoplast intrinsic protein) subfamily. TIP is
an integral membrane protein that was originally found in plant
seeds and belongs to the MIP (major intrinsic protein) family,
the members of which are widely distributed in bacteria, animals
and plants. The TIP of plant seeds is abundant and is conserved
among both monocots and dicots. The predicted 29-kDa precursor
to MP23 includes six putative membrane-spanning domains, and the
first loop between the first and second transmembrane domains
is larger than that of MP28. The N-terminal sequence of the mature
MP23 starts from residue 66 in the first loop, indicating that
an N-terminal 7-kDa fragment that contains one transmembrane domain
is post-translationally removed. During maturation of pumpkin
seeds, mRNAs for MP28 and MP23 became detectable in cotyledons
at the early stage, and their levels increased slightly until
a rapid decrease occurred at the late stage. This is consistent
with the accumulation of the 29-kDa precursor and MP28 in the
cotyledons at the early stage. By contrast, MP23 appeared at the
late stage simultaneously with the disappearance of the 29-kDa
precursor. Thus, it seems possible that the conversion of the
29-kDa precursor to the mature MP23 might occur in the vacuoles
after the middle stage of seed maturation. Both proteins were
localized immunocytochemically on the membranes of the vacuoles
at the middle stage and the protein bodies at the late stage.
These results suggest that both MP28 and the precursor to MP23
accumulate on vacuolar membranes before the deposition of storage
proteins, and then the precursor is converted to the mature MP23
at the late stage. These two TIPs might have a specific function
during the maturation of pumpkin seeds.
Processing enzymes responsible for the maturation of seed proteins
belong to a novel group of cysteine proteinases with molecular
masses of 37 to 39 kDa. However, the processing enzyme activity
can be found not only in seeds but also in vegetative tissues
such as hypocotyls, roots and mature leaves. Thus, we designated
the enzyme as a vacuolar processing enzyme (VPE). The molecular
characterization of all the members of the VPE family in Arabidopsis
is required if we are to elucidate the mechanisms of regulation
of genes for VPE homologues and the physiological functions of
these proteins in protein-storage vacuoles and vegetative vacuoles.
Southern blot analysis showed that a family of genes for VPEs
in Arabidopsis thaliana was composed of three genes, for
Alpha-VPE, Beta-VPE and Gamma-VPE. We isolated the three genes
of VPEs from a genomic library of Arabidopsis. The positions
of eight introns were fully conserved among the three genes, with
the exception that the Alpha-VPE gene was missing the fifth intron
found in the Beta-VPE and Gamma-VPE genes.
Northern blot analysis revealed that Alpha-VPE was expressed in
rosette leaves, cauline leaves and stems of Arabidopsis,
while Beta-VPE was predominantly expressed in the flowers and
buds. The Gamma-VPE gene was expressed predominantly in the stems,
with a lower level of expression in rosette and cauline leaves.
However, the expression was not detected in roots, flowers plus
buds, or green siliques. This result strongly suggests that the
Alpha-VPE and Gamma-VPE genes encode isozymes of VPE that are
specific to vegetative organs. To demonstrate temporal and spacial
expression of the promoters of the vegetative VPE gene, we transformed
tobacco plants with a reporter gene containing the promoter of
the Gamma-VPE gene and the coding region of Beta-glucuronidase
(GUS). The GUS activity was predominantly expressed in the senescing
tissues (Fig. 2).
Members of the VPE family can be separated into two subfamilies,
one that is specific to seeds and another that is specific to
vegetative organs, such as leaves and stems. The members of the
seed subfamily might function in the protein-storage vacuoles
of seeds, while those of the vegetative subfamily might function
in the lytic vacuoles of non-storage organs. The VPE cleaved a
peptide bond on the carbonyl side of an exposed asparagine residue
on the molecular surface of proprotein precursors to generate
the mature forms of seed proteins. A similar type of post-translational
processing has been reported in the case of maturation of vacuolar
proteins in vegetative tissues, such as the proteinase inhibitors
of tomato leaves and tobacco stigmas, and the chitinase of tobacco
leaves and cultured cells. These observations suggested that VPEs
are widely distributed in plant tissues and play crucial roles
in the maturation of a variety of proteins in plant vacuoles.
This suggestion is supported by the report that VPE activity can
be detected not only in seeds but also in vegetative organs and,
moreover, that two of the VPE genes of Arabidopsis are
specifically expressed in vegetative organs.

Publication List:
De Bellis, L., Hayashi, M., Nishimura, M. and Alpi, A. (1995) Subcellular distribution of aconitase isoforms in pumpkin cotyledons. Planta 195, 464-468.
Hara-Nishimura, I., Shimada, T., Hiraiwa, N. and Nishimura, M. (1995) Vacuolar processing enzyme responsible for maturation of seed proteins. J. Plant Physiol. 145, 632-640.
Hayashi, M., De Bellis, L., Alpi, A. and Nishimura, M. (1995) Cytosolic aconitase is a member for glyoxylate cycle in etiolated pumpkin cotyledons. Plant Cell Physiol. 36, 669-680.
Inoue, K., Motozaki, A., Takeuchi, Y,. Nishimura, M. and Hara-Nishimura, I. (1995) Molecular characterization of proteins in protein-body membrane that disappear most rapidly during transformation of protein bodies into a vacuole. Plant Journal 7, 235-243.
Inoue, K., Takeuchi, Y., Nishimura, M. and Hara-Nishimura, I. (1995) Characterization of two integral membrane proteins located in the protein bodies of pumpkin seeds. Plant Mol. Biol. 28, 1089-1101.
Kato, A., Hayashi, M., Mori, H. and Nishimura, M. (1995) Molecular characterization of a glyoxysomal citrate synthase that is synthesized as a precursor of higher molecular mass in pumpkin. Plant Mol. Biol. 27, 377-390.
Kinoshita, T., Nishimura, M. and Hara-Nishimura, I. (1995) Homologues of a vacuolar processing enzyme that are expressed in different organs in Arabidopsis thaliana. Plant Mol. Biol. 29, 81-89.
Kinoshita, T., Nishimura, M. and Hara-Nishimura, I. (1995) A family of the vacuolar processing enzymes in Arabidopsis thaliana is encoded by three genes: The sequence and expression of the third g-VPE gene. Plant Cell Physiol. 36, 1555-1562.
Nii, N., Yamaguchi, K. and Nishimura, M. (1995) Effects of fruiting on amylase activity and ribulose bisphosphate carboxylase-oxygenase content in peach leaves. J. Jap. Soc. Hort. Sci. 64, 267-273.
Nii, N., Watanabe, T., Yamaguchi, K. and Nishimura, M. (1995) Changes of anatomical features, photosynthesis and ribulose bisphosphate carboxylase-oxygenase content of mango leaves. Annals of Botany 76, 649-656.
Nozue, M., Kubo, H., Nishimura, M. and Yasuda, H. (1995) Detection and characterization of a vacuolar protein (VP24) in anthocyanin-producing cells of sweet potato suspension cultures. Plant Cell Physiol. 36, 883-889.
Strzalka, K., Hara-Nishimura, I. and Nishimura, M. (1995) Physical change in vacuolar membranes during transformation of protein bodies into vacuoles in germinating pumpkin seeds. Biochim. Biophys. Acta 1239, 103-110.
Takeda, S., Kowyama, Y., Takeuchi, Y., Matsuoka, K., Nishimura, M. and Nakamura, K. (1995) Spatial patterns of sucrose-inducible and polygalacturonic acid-inducible expression of genes that encode sporamin and b-amylase in sweet potato. Plant Cell Physiol. 36, 321-333.
Yamaguchi, K., Mori, H. and Nishimura, M. (1995) A novel isoenzyme of ascorbate peroxidase localized on glyoxysomal and leaf peroxisomal membranes in pumpkin. Plant Cell Physiol. 36, 1157-1162.
Yamaguchi, K., Takeuchi, Y., Mori, H. and Nishimura, M. (1995) Development of microbody membrane proteins during the transformation of glyoxysomes to leaf peroxisomes in pumpkin cotyledons. Plant Cell Physiol. 36, 455-464.
The Division has been closed upon Professor Yoshihiko Fujita's retirement on March 31, 1995, and will be re-intiated in 1996 on new projects.
Fujita, Y., Murakami, A., and Aizawa, K. (1995) The accumulation of protochlorophilide in cells of Synechocystis PCC 6714 with a low PSI/PSII stoichiometry. Plant Cell Physiol. 36, 575-582.
Ohki, K. and Fujita, Y. (1995) Intracellular location of cytochrome oxidase active in vivo in the cyanophytes, Synechocystis sp. PCC 6714 and Anacystis nidulans Tx20 and R2, grown under various conditions. Protoplasma 188, 70-77.
Professor: Masayuki Yamamoto
Research Associate: Masuo Goto
NIBB Postdoctoral Fellow: Takashi Kuromori
Graduate Students: Satsuki Okamoto (from Nara Women's University),
Fumihiko Hakuno (from The University of Tokyo), Chikako Kitayama
(from The University of Tokyo)
This Division aims to explore the regulation of meiosis in higher
organisms. Meiosis is a crucial step in gamete formation and is
essential for sexual reproduction. Meiotic steps are highly conserved
among eukaryotic species. The major strategy taken by us to isolate
genes that may be relevant to the regulation of meiosis in animals
or plants depends upon trans-complementation between heterologous
organisms. In the fission yeast Schizosaccharomyces pombe,
which is a unicellular eukaryotic microorganism, genes involved
in control of meiosis have been well characterized and many of
them are cloned (reviewed in Yamamoto (1996) Trends Biochem. Sci.
21, 18-22). Mutants defective in these genes, isolated either
by classical genetics or by gene disruption and chromosome manipulation,
are available. We have thus set out to isolate homologs of these
S. pombe genes from animals and a plant, mainly by using
functional complementation of the mutants. To facilitate this
strategy, we also paid efforts to elucidation of the regulatory
mechanisms of meiosis in the fission yeast.
Using cDNA libraries prepared from mouse testis, Xenopus
oocyte and Arabidopsis thaliana, we screened extensively
for genes that can rescue loss of function of the following three
genes, which are involved in the regulation of sexual development
in S. pombe: The sme2 gene, which encodes an RNA
product essential for the promotion of meiosis I; the pde1
gene, which encodes cAMP phosphodiesterase; and the mes1
gene, which is required for the promotion of meiosis II. Genes
encoding putative kinases, transcription factors, RNA-binding
proteins and others have been isolated in these screenings, and
their possible roles in the regulation of meiosis are currently
under investigation. In particular, we found that various genes
encoding cytoskeletal proteins can suppress the mes1 defect,
suggesting that the regulation of meiosis II is closely related
to modification or reorganization of the cytoskeleton.
The cdc2 gene of S. pombe encodes a serine/threonine protein kinase, which plays key roles in the progression of both G1 and G2 phases in the mitotic cell cycle. This kinase regulates G2/M transition in cooperation with a B-type cyclin encoded by cdc13, forming a complex called MPF (M-phase promoting factor). The cdc25 gene encodes a tyrosine protein phosphatase that activates cdc2 kinase by dephosphorylation. These three cdc genes are required for mitosis, and cells carrying a temperature-sensitive mutation in either of these genes frequently produce two-spored asci at the semi-restrictive temperature, apparently not undergoing meiosis II. Cells carrying a mutant allele of cdc2, originally named tws1, have no obvious defect in the mitotic cell cycle but produce two-spored asci skipping meiosis II. These previous observations suggest that these three genes are essential for meiosis II. However, the involvement of these genes in meiosis I has been left unclear. We carefully investigated this problem in three different experimental systems and concluded that the function of cdc2 is essential for premeiotic DNA synthesis, and that cdc13 and cdc25 are essential for meiosis I. We could not conclude whether cdc2 is essential for meiosis I, due to experimental difficulties, although it appears likely. The necessity of cdc2 function for premeiotic DNA synthesis in S. pombe contrasts to the previous report by others that CDC28, the S. cerevisiae homolog of cdc2, is dispensable for it.

Iino, Y., Hiramine, Y. and Yamamoto, M. (1995) The role of cdc2 and other genes in meiosis in Schizosaccharomyces pombe. Genetics 140, 1235-1245.
Izumiya, H. and Yamamoto, M. (1995) Cloning and functional analysis of the ndk1 gene encoding nucleoside diphosphate kinase in Schizo-saccharomyces pombe. J. Biol. Chem. 17, 27859-27864.
Kanoh, J., Sugimoto, A. and Yamamoto, M. (1995) Schizosaccharomyces pombe zfs1+ encoding a zinc-finger protein functions in the mating pheromone recognition pathway. Mol. Biol. Cell 6, 1185-1195.
Professor: Yoshiki Hotta
Associate Professor: Hitoshi Okamoto
Research Associates: Mika Tokumoto, Shin-ichi Higashijima
Institute Research Fellow: Nobuyoshi Shimoda
Brain can be seen as an integrated circuit where neurons of various identities are interconnected in a highly ordered manner by their axons. We have been interested in how individual neurons aquire their own identities and how their axons find their own pathways and finally recognize their proper targets. Using zebrafish (Danio rerio), which is suitable for genetic analysis and gene manipulation, we are trying to address these questions both at the molecular and cellular levels.
Islet-1 (Isl-1) is a LIM domain/homeodomain-type transcription
regulator originally identified as an insulin gene enhancer binding
protein. Isl-1 is considered to be involved in the differentiation
of the neuronal cells. We previously cloned a isl-1 homolog
from zebrafish cDNA library, named it zebrafishisl-1. Recently,
we have isolated two novel isl-1 homologs from zebrafish
cDNA library, named them zebrafishisl-2 and 3.
We examined the mRNA expression pattern of each homolog using
in situ hybridization to whole-mount embryos. All three homologs
are expressed in Rohon-Beard neurons. However the expression in
primary motoneurons diverged. Zebrafishisl-1 mRNA is expressed
in the rostral primary motoneuron (RoP). isl-2 mRNA is
expressed in the caudal primary motoneuron (CaP) and its variant
sibling (VaP). isl-3 mRNA is expressed in the ventral region
of the myotome but not in the primary motoneurons. The ventral
myotome is the region that the axon of the CaP extend into. isl-3
mRNA is also expressed throughout the developing eye and tectal
region of the midbrain, the target for the retial axons. These
results raise possibilities that the isl-1 homologs may
be involved in the specification and/or target recognition by
the primary motoneurons.
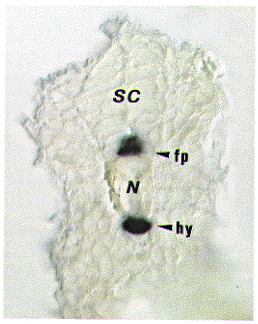
F-Spondin is a secreted protein expressed at high levels in the floor plate. The C-terminal half of the protein contains six TSRs (Thrombospondin Type 1 repeats), while the N-terminal half exhibited no homology to other proteins. Functions of F-Spondin in vivo remain largely unknown. AN34, expressed in a subset of muscle cells in Drosophila, was cloned and found to encode a secreted protein sharing high degree of homology in two parts (H1 and H2) with the N-terminal half of F-Spondin (personal communication). This suggests that the homologous reigions are novel domains important for neuronal development. Thus, we searched for AN34/F-Spondin family from zebrafish (Danio rerio). By PCR screening and subsequent cDNA screening, we identified two novel genes (AN1 and AN2) in addition to zebrafish F-Spondin. AN1 and AN2 consist of H1, H2 and one TSR. The overall structure are highly similar to AN34 and more related to AN34 than F-Spondin, suggesing that AN1, AN2 and Drosophila AN34 constitute a novel subfamily. AN1 is expressed weakly in the floor plate and a small subset of neuronal cells at 22-26 hr. AN2 expression is first observed broadly around the axial mesoderm (notochord) at 10-12 hr. Then, the expression becomes restricted to the floor plate and hypochord at 14 hr and continues at high levels up to 26 hr. F-Spondin is expressed in the floor plate as seen in rat. These findings suggest that the AN34/F-Spondin family play important roles in the midline development in the vertebrate.
We have discovered two families of short interspersed repetitive
elements (SINEs) in the zebrafish genome. The two families are
marked by their wide distribution: one family, designated mermaid,
is also present in the genomes of other fish, amphibian and primates,
but absent in the mouse genome. Some members of the mermaid
family were found in transposon-like repetitive elements including
Tc1-like elements which are also distributed in the genomes of
fish and amphibian. This raises the possibility of horizontal
transfer of the mermaid family between vertebrates via
DNA-mediated transposition. The mermaid family is distinctive
in each species except for a conserved region of approximately
80 bp. The zebrafish mermaid sequence is about 400 bp long
and has a typical SINE structure: split promoter of RNA polymerase
III at the 5' end, tandem repeats of short oligonucleotide (AATG)n
at the 3' end and target site duplications.
The zebrafish mermaid were estimated to be 12,000 copies
per haploid genome and highly polymorphic between AB comp and
Darjeeling strains. We also found that oligonucleotides directed
to the conserved region of the mermaid family can be used
to recover zebrafish specific DNA from zebrafish-mouse cell hybrids
by PCR (mermaid PCR). Thus, the mermaid sequence
serves as a valuable genetic tool for the zebrafish genome mapping.
Another SINE family is distributed not only in other Cypriniformes,
such as silver carp and grass carp but also in medaka, at least.
The family was named angel since some DNA fragments cloned
from medaka which harbor the entire region of the SINE migrated anomalously on gels. The anomaly was temperature-dependent, suggesting
a sequence-induced unusual structure of DNA. The angel
family belongs to a new class of elements referred to as miniature
inverted repeat transposable elements (MITEs) or inverted repeat
SINEs. The angel sequence has relatively long terminal
inverted repeats (TIRs) that flank short non-coding DNA. The TIRs
of angel begin with the sequence TTAAAGGRR, known as the
T2 motif first identified in other inverted repeat SINEs in Xenopus.
Chiba, A., Snow, P., Keshishian, H. and Hotta, Y. (1995). Fasciclin III as a synaptic target recognition molecule in Drosophila. Nature 374, 166-168.
Hosoya, T., Takizawa, K., Nitta, K. and Hotta, Y. (1995). glial cells missing: A binary switch between neuronal and glial determination in Drosophila. Cell 82, 1025-1036.
Tokumoto, M., Gong, Z., Tsubokawa, T., Hew, C. L., Uyemura, K., Hotta, Y. and Okamoto, H. (1995). Molecular heterogeneity among primary motoneurons and within myotomes revealed by the differential mRNA expression of novel Islet-1 homologs in embryonic zebrafish. Dev. Biol. 171, 578-589.
Professor: Hitoshi Sakano
Research Associate: Masahiro Ishiura (until March), Fumikiyo
Nagawa (until April), Kanae Muraiso (on leave), Akio Tsuboi
Institute Research Fellow: Hiroaki Kasai
Graduate Student: Setsuyuki Aoki (from Kyoto University), Nika
Yamazaki (from Tokyo Institute of Technology)
Our research interest has been focused on somatic DNA changes
in the immune system. In lymphocytes, DNA recombination and gene
conversion play important roles in the expression of antigen receptor
genes. Gene rearrangement, known as V-(D)-J joining, not only
generates a vast diversity in the receptor genes, but also insures
the activation of a particular member of the multigene family.
Recently, hundreds of odorant receptor (OR) genes have been reported
in the olfactory system, although it is yet to be studied how
this gene system is regulated for expression. Each OR gene is
expressed in one of the four topographically distinct zones, where
olfactory neurons expressing a given OR gene are randomly distributed.
It is assumed that only a limited number of the OR genes (possibly
one) are activated in each olfactory neuron.
We have been studying how individual neurons express a limited
number of OR genes keeping the rest of the genes silent. In order
to study this selective expression of the OR genes, we have been
characterizing a P1 clone containing two highly related genes,
MOR10 and MOR28. These genes are 92% homologous in the coding
regions and linked in tandem on mouse chromosome 14. In situ
hybridization has revealed that both genes are expressed in the
same spatial zone within the olfactory epithelium, but never expressed
simultaneously in the same neuron. Interestingly, olfactory neurons
expressing either MOR10 or MOR28 project their axons to a distinct,
but adjacent set of glomeruli on the olfactory bulb.
Developmental analyses have revealed that MOR10-expressing neurons
come out earlier than MOR28-expressing ones during mouse embryogenesis.
However, the number of MOR28 neurons overcomes that of MOR10 neurons
after birth; the ratio of cells expressing MOR28 vs. MOR10 reaches
2.5 : 1 in the adult. These results suggest that individual olfactory
neurons may activate OR genes through a stochastic mechanism even
between two neighboring and closely related OR genes; yet this
selection appears to be biased in both the onset and levels of
expression.
In order to study whether the projection of axons to the olfactory
bulb exerts influences on the OR gene expression, we have characterized
pdn/pdn mutant mice in collaboration with Dr. Naruse at
Aichi prefectural colony. The homozygous mice show a number of
developmental abnormalities including polydactyly and gross malformations
of the brain; they also lack olfactory bulbs. In situ hybridization
has revealed that expression patterns of the MOR10 and MOR28 genes
in the pdn/pdn embryos are comparable to those found in
the wild-type, suggesting that the zonal expression of OR genes
in the olfactory epithelium is regulated independently of influences
from the olfactory bulb.
For the study of the mutually exclusive expression of OR genes,
we have generated transgenic mice which are devised to express
the MOR28 gene in every olfactory neuron under the control of
the olfactory marker protein (OMP) promoter. Since the OMP gene
is expressed in the mature neurons within the olfactory epithelium,
the transgene is expected to be activated in all olfactory neurons.
We believe that the study of such transgenic mice will give us
new insight into the molecular mechanisms for OR gene expression,
as well as for neuronal projection to the olfactory bulb.
When the olfactory neurons are regenerated, they send axons to
specific sets of glomeruli in the olfactory bulb. It is amazing
that axons expressing a given OR gene are able to find their target
among two thousand pairs of glomeruli. We hope that our transgenic
approach mentioned above will become a useful clue to study the
target specificity and selectivity in the synapse formation.
Aoki, S., Kondo, T. and Ishiura, M. (1995) Circadian expression of the dnaK gene in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 177, 5606-5611.
Hiramatsu, R., Akagi, K., Matsuoka, M., Sakumi, K., Nakamura, H., Kingsbury, L., David, C., Hardy, R. R., Yamamura, K. and Sakano, H. (1995) The 3' enhancer region determines the B/T specificity and pro-B/pre-B specificity of immunoglobulin VKappa-JKappa joining. Cell 83, 1113-1123.
Liu, Y., Golden, S. S., Kondo, T., Ishiura, M. and Johnson, C. H. (1995) Bacterial luciferase as a reporter of circadian gene expression in cyanobacteria. J. Bacteriol. 177, 2080-2086.
Liu, Y., Tsinoremas, N. F., Johnson, C. H., Lebedeva, N. V., Golden, S. S., Ishiura, M. and Kondo, T. (1995) Circadian orchestration of gene expression in cyanobacteria. Genes Dev. 9, 1469-1478.
Moriya, H., Kasai, H. and Isono, K. (1995) Cloning and characterization of the hrpA gene in the terC region of Escherichia coli that is highly similar to the DEAH family RNA helicase genes of Saccharomyces cerevisiae. Nucleic Acids Res., 23, 595-598.