派生形質
導管
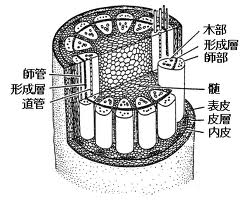
維管束は、植物がもつ内部組織の1つで、、水分や養分(無機塩類)の通路である木部 xylem と 光合成産物などの栄養分の通路である師部からなる。
木部は、導管、仮導管、木部放射細胞、木部柔細胞、木部繊維からなる。
一方、師部は、師管、伴細胞、師部放射細胞、師部柔細胞、師部繊維からなる。道管は完成すると細胞が死んで細胞質がなくなり、後に水を運ぶ役割を果たすから、木部の大部分は死んだ組織である。
なお、師部と木部の間には、分裂組織である維管束形成層が存在する場合がある。
仮導管
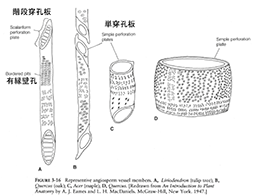
シダ植物と一部の被子植物(アンボレラ科)、一部の裸子植物で見られる導管は、境界の細胞壁が消失していない仮導管である。これらの植物では、仮道管が水の通道と機械的支持の両機能を果たしている。仮道管は大変細長い細胞で、両端は次第にとがり、隣接する細胞と連結している。細胞が成熟するにつれ、一次壁上に二次壁物質(リグニン)が蓄積し壁が肥厚すると共に、中の細胞質は失われていく。二次壁には、二次肥厚の認められない縁取りのある穴(有縁壁孔bordered pits)が多数あり、この壁孔を通して水の通道が行われる。
モクレン亜網のシキミモドキ科、アンボレラ科、マンサク亜網のヤマグルマ科やスイセイジュ科、そしてスイレンを中心とする多くの水生植物などは、無道管被子植物として知られている。
花粉

花粉(かふん)とは、種子植物門の植物の花の雄蘂(おしべ)から出る粉状の細胞。花粉がめしべの先端(柱頭)につくことにより受粉が行われる花粉は細胞壁が厚くなり、形は種によって異なり、表面にはそれぞれの種で特有の構造を持つ。
真正双子葉類は三溝粒 Tricolpates の花粉をもつ。これは花粉の発芽溝または発芽孔が基本的に3個あるという意味である(二次的に数が変わったものもある)。一方、マツモ科、センリョウ科、モクレン目、クスノキ目、コショウ目、カネラ目、そして単子葉類はこれは基本的に1個の単溝粒の花粉を形成する。この形質は裸子植物とも共通で、共有原始形質と考えられる。1990年代には分子系統学の立場からも、この考えが支持されるようになった。

テクタム
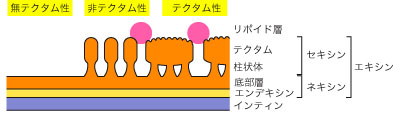
花粉粒の細胞壁は、花粉壁 (sporoderm) とよばれる。花粉壁は左図のように複雑な層状構造を示す(図参照)。
柱状体 (柱状層、円柱体) (columella, bacula) :セキシンの内層であり、底部層の表面に直立した柱状の構造。花粉壁において柱状体を欠くものを無テクタム性 (atectate) という (例:アンボレラ科、モクレン科、バンレイシ科) 。
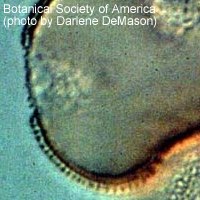
テクタム (tectum 外表層) :柱状体の頭部が融合したもの。
全てが切れ目無く融合して花粉粒を覆っていることもあるし (外表層型) 、一部に孔の開いた状態で取り残されて網目状になるものもある (半外表層型) 。
柱状体が独立しておりテクタムが形成されないものを非テクタム�ォ (intectate) という (外表層欠失型) 。
それに対してテクタムをもつものをテクタム性 (tectate) という。
心皮
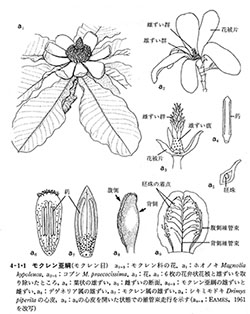
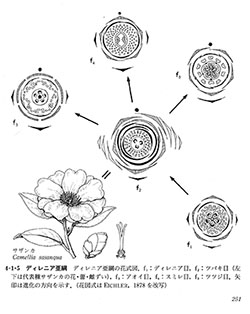
心皮は胚珠を包み込む雌性の花器官であり、胚珠は心皮の壁で囲われた部屋の中に閉じ込められている。一つの花に心皮が多くあるもの(多心皮)から1個の場合まである。
複数の心皮が離れた状態を離生心皮(図4-1-1, a3)、心皮が隣同士癒合してできたものを合生心皮(図4-1-5, f2)という。合生心皮では部屋の数は元の心皮数と同じであるが、減少することも多い。
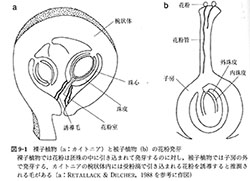
心皮はとっくり状で、花粉が発芽する柱頭、花粉管が中を伸長する棒状の花柱、および部屋があって胚珠を内包する子房からなる(図9-1)。原始的な心皮では部屋が心皮の大部分を占め、柱頭も心皮の先端から中下部まで伸びて、各部分が区別しにくい(図4-1-1, a8)。(植物の多様性と系統; 岩槻邦男・馬渡峻輔 監修, 裳華房 より)

 シキミ科では、花被は螺旋状、雄蕊や雌蕊は輪状に配列しており、何によって螺旋から輪ができるのかの研究に適しているかもしれない。
シキミ科では、花被は螺旋状、雄蕊や雌蕊は輪状に配列しており、何によって螺旋から輪ができるのかの研究に適しているかもしれない。