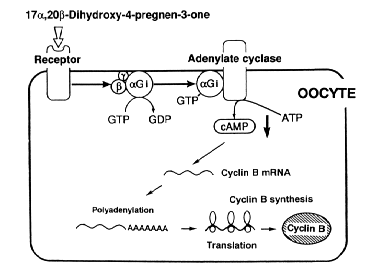
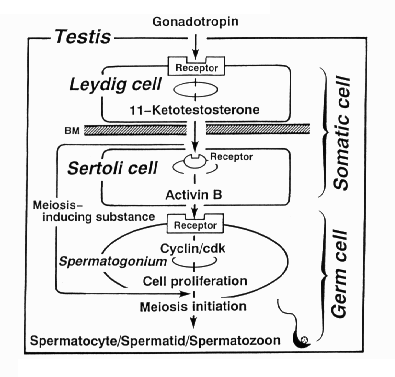
NATIONAL INSITUTE FOR BASIC BIOLOGY
National Institute for Basic Biology
Division of Reproductive Biology
- Professor:
- Yoshitaka Nagahama
- Associate Professor:
- Michiyasu Yoshikuni
- Research Associates:
- Minoru Tanaka
- Tohru Kobayashi
- Institute Research Fellow:
- Akihiko Yamaguchi
- JSPS Postdoctoral Fellows:
- Yoshinao Katsu
- Zuxu Yao
- NIBB Postdoctoral Fellows:
- Kunimasa Suzuki
- Won-Kyo Lee
- Jain-Quiao Jiang
- Xiao-Tian Chang
- Won-Kyo Lee
- JSPS Research Associates:
- Mika Takahashi
- Takashi Todo
- Daisuke Kobayashi
- Takashi Todo
- Graduate Students:
- Yuichi Ohba (Graduate University for Advanced Studies)
- Jun Ding (Graduate University for Advanced Studies)
- Masatada Watanabe (Graduate University for Advanced Studies)
- Guijun Guan (Graduate University for Advanced Studies)
- Jun Ding (Graduate University for Advanced Studies)
- Monbusho Foreign Scientist:
- Graham Young (University of Otago)
- JSPS Visiting Scientist:
- Allen Schuetz (University of Maryland)
- Visiting Scientist:
- Craig Morrey (University of Hawaii)