Three regular divisions and one adjunct division that has been reinitiated on new projects during 1995, construct the Department. The department members conduct molecular analysis on various aspects of developmental phenomena; differentiation and maturation of the germ cells, molecular basis of the body plans, and gene regulation in different phases of the cell cycle, in the differentiating tissues, and during changes of the differentiated states.
Professor: Yoshitaka Nagahama
Associate Professor: Michiyasu Yoshikuni
Research Associates: Minoru Tanaka, Tohru Kobayashi
JSPS Postdoctoral Fellow: Yoshinao Katsu
Postdoctoral Fellows: Wei Ge, Ahn Ryun-Sap, Kazumasa Suzuki,
Takashi Todo
Graduate Students: Shinji Onoe (Graduate University for Advanced
Studies), Xiao-Tian Chang (Graduate University for Advanced Studies),
Daisuke Kobayashi (Graduate University for Advanced Studies),
Yuichi Ohba (Graduate University for Advanced Studies), Jun Ding
(Graduate University for Advanced Studies), Masatada Watanabe
(Graduate University for Advanced Studies), Shigenari Ijiri (Hokkaido
University)
Visiting Scientists: Joel Sohn (California Institute of Technology),
Craig Morrey (University of Hawaii)
Jian-Quiao Jiang (University of Wuhan)
The division of reproductive biology conducts research on the endocrine regulation of differentiation, growth and maturation of germ cells in multicellular animals, using fish as a primary study model.
Our research effort in previous years concentrated on the identification
and characterization of the molecules (pituitary gonadotropins
and gonadal steroid hormones) that stimulate and control germ
cell growth and maturation. It was in 1985 that we identified,
for the first time in any vertebrate, 17Alpha, 20Beta-dihydroxy-4-pregnen-3-one
(17Alpha, 20Beta-DP) as the maturation-inducing hormone of amago
salmon (Oncorhynchus rhodurus). Along with estradiol-17Beta,
which was identified as the major mediator of oocyte growth, we
now have two known biologically important mediators of oocyte
growth and maturation in female salmonid fishes. It is established
that the granulosa cells are the site of production of these two
mediators, but their production by the ovarian follicle depends
on the provision of precursor steroids by the thecal cell (two-cell
type model). A dramatic switch in the steroidogenic pathway from
estradiol-17Beta to 17Alpha, 20Beta-DP occurs in ovarian follicle
cells immediately prior to oocyte maturation. This switch is a
prerequisite step for the growing oocyte to enter the maturation
phase, and requires a complex and integrated network of gene regulation
involving cell-specificity, hormonal regulation, and developmental
patterning.
We have isolated and characterized cDNAs encoding several ovarian
steroidogenic enzymes of rainbow trout (Oncorhynchus mykiss),
medaka (Oryzias latipes), tilapia (Oreochromis niloticus),
and ayu (Plecoglossus altivelis) which are responsible
for estradiol-17Beta and 17Alpha, 20Beta-DP biosynthesis: cholesterol
side-chain cleavage cytochrome P450 (P450scc), 3Beta-hydroxysteroid
dehydrogenase (3Beta-HSD), 17Alpha-hydroxylase/C17,20-lyase cytochrome
P450 (P450c17), cytochrome P450 aromatase (P450arom) and 20Beta-hydroxysteroid
dehydrogenase (20Beta-HSD). We also isolated the structural genes
encoding P450c17 and P450arom from medaka. The medaka P450arom
gene consists of nine exons, but spans only 2.6 kb, being much
smaller than the human P450arom gene (at least 70 kb), as the
result of extremely small introns.
Northern blots using a tilapia P450arom cDNA fragment and Western
blots using an antiserum against a tilapia P450arom oligopeptide
fragment revealed a single P450arom mRNA (2.6 kb) and a single
protein (59 kDa) in tilapia ovarian tissue, respectively. These
analyses also reveal that the levels of both P450arom mRNA and
protein are low in early vitellogenic follicles, increase in mid-vitellogenic
follicles, and decline to non-detectable levels in post-vitellogenic
follicles. Changes in the ability of follicles to convert exogenous
testosterone to estrogens (aromatase activity) are similar to
those of P450arom mRNA and protein. These observations indicate
that the capacity of tilapia ovarian follicles to synthesize estradiol-17Beta
is closely related to the contents of P450arom mRNA and protein
within them.
In rainbow trout and medaka, P450scc and P450c17 (also 3Beta-HSD
in rainbow trout) mRNA transcripts increase in follicles towards
the end of oocyte growth phase and during oocyte maturation. Furthermore,
incubations of isolated thecal layers with gonadotropin result
in the elevation of P450scc mRNA. The effect of gonadotropin becomes
more dramatic when the expression of P450scc mRNA is examined
in granulosa cells. P450scc mRNA is not detected in the absence
of gonadotropin, but markedly expresses in the presence of gonadotropin.
The increase in the amount of P450scc, 3Beta-HSD and P450c17 transcripts
provide an explanation for the dramatic increase in 17Alpha, 20Beta-DP
production in follicles during naturally- and gonadotropin-induced
oocyte maturation. In contrast, levels of mRNA for P450arom are
high during oocyte growth, but rapidly decrease during oocyte
maturation. This decrease in P450arom mRNA levels appears to be
correlated with the decreased ability of maturing follicles to
produce estradiol-17Beta.
17Alpha, 20Beta-DP acts via a receptor on the plasma membrane
of oocytes. We have identified and characterized a specific 17Alpha,
20Beta-DP receptor from defolliculated oocytes of several fish
species. 17Alpha,
20Beta-DP receptor concentrations increase during oocyte maturation.
Pertussis toxin (PT) catalyzed the ADP ribosylation of a 40 kDa
protein in crude membranes from rainbow trout oocytes. The 40
kDa protein is recognized by an antibody against a subunit of
inhibitory G-protein. Treatment of the membrane fraction with
17Alpha, 20Beta-DP decreases the PT-catalyzed ADP ribosylation
of the 40 kDa protein. The specific binding of 17Alpha, 20Beta-DP
is decreased by PT. We conclude that the PT-sensitive Gi is involved
in the signal transduction pathway of 17Alpha, 20Beta-DP in fish
oocytes.
The early steps following 17Alpha, 20Beta-DP action involve the
formation of the major mediator of this steroid, maturation-promoing
factor or metaphase-promoting factor (MPF). MPF activity cycles
during 17Alpha, 20Beta-DP. induced oocyte maturation with the
highest activity occurring at the first and second meiotic metaphase.
Studies from our laboratory and others have shown that MPF activity
is not species-specific and can be detected in both meiotic and
mitotic cells of various organisms, from yeast to mammals. Fish
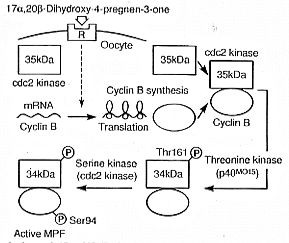
MPF, like that of amphibians, consists of two components, catalytic
cdc2 kinase (34 kDa) and regulatory cyclin B (46 to 48 kDa).
Immature goldfish oocytes contain only monomeric 35 kDa cdc2.
Although immature oocytes contain mRNA for cyclin B, they do not
contain cyclin B protein. 17Alpha, 20Beta-DP induces oocytes to
synthesize cyclin B. The preexisting 35 kDa inactive cdc2 kinase
binds to de novo synthesized cyclin B at first, and then
is rapidly converted into the 34 kDa active form by a threonine
(Thr) kinase, p40MO15 (Fig.1).
Introduction of a bacterially-produced goldfish cyclin B into
immature goldfish oocyte extracts induces cdc2 kinase activation.
Phosphoamino acid analysis shows that Thr phosphorylation of the
34 kDa cdc2 kinase is associated with the activation. The site
of Thr phosphorylation on cdc2 kinase was mapped to residue Thr
161. Since goldfish cyclin B mRNA contains four copies of the
usual cytoplasmic polyadenylation element in the 3'UTR, the initiation
of its synthesis during oocyte maturation may be controlled by
the elongation of poly (A) tail, as suggested in other systems.
We exmined the polyadenylation state of cyclin B mRNA during goldfish
oocyte maturation by means of a PCR poly (A) test, and found that
cyclin B mRNA is polyadenylated during oocyte maturation.
Immediately prior to the transition from metaphase to anaphase,
M-phase-promoting factor (MPF) is inactivated by degradation of
cyclin B. We investigated the role of proteasomes (a nonlysosomal
large protease) in cyclin degradation, using Eschelicia coli-produced
goldfish cyclin B and purified goldfish proteasomes (20S and 26S).
The purified 26S proteasome, but not 20S proteasome, cleaved both
monomeric and cdc2-bound cyclin B at lysine 57 (K57) restrictively
in vitro, and produced a 42 kDa N-terminal truncated cyclin
B, which was transiently detected at the initial phase of the
normal egg activation. The 42 kDa cyclin B, as well as full-length
one, was degraded in Xenopus egg extracts, but a mutation
on K57 (K57R) inhibited both the digestion by 26S proteasome and
the degradation in Xenopus egg extracts. These findings
strongly suggest the involvement of 26S proteasome in cyclin degradation
through the first cleave on its N-terminus.
We have identified two steroidal mediators of male germ cell development
in salmonid fishes (11-ketotestosterone for spermatogenesis and
17Alpha, 20Beta-DP for sperm maturation). 11-ketotestosterone
can also induce the entire process of spermatogenesis in an organ
culture system developed for eel testes from premitotic spermatogonia
to spermatozoa within 21 days. A steroidogenic switch, from 11-ketotestosterone
to 17Alpha, 20Beta-DP, occurs in salmonid testes around the onset
of final maturation. In vitro incubation studies using
different testicular preparations have revealed that the site
of 17, 20-DP production is in the sperm, but its production depends
on the provision of precursor steroids by somatic cells. The site
of 11-ketotestosterone production is in the testicular somatic
cells.
In the cultivated male Japanese eel (Auguilla japonica),
spermatogonia are the only germ cells present in the testis. Subtractive
hybridization was used to identify genes that are expressed differentially
in eel testes in the first 24 hr after HCG treatment in vivo,
which ultimately induces spermatogenesis. One up-regulated cDNA
was isolated from subtractive cDNA libraries derived from mRNA
extracted from control testes and testes one day after a single
injection of HCG. From its deduced amino acid sequence, this clone
was identified as coding for the activin B subunit. The HCG-dependent
activin BetaB mRNA expression in the testes was restricted to
Sertoli cells in testes treated with HCG for one to three days.
A marked stimulation of activin B production was observed in testes
after HCG and 11-ketotestosterone treatment. Addition of recombinant
human activin B induced spermatogonial proliferation in vitro.
Taken together, these findings suggest the following sequence
of the hormonal induction of spermatogenesis in the eel. Gonadotropin
stimulates the Leydig cells to produce 11-ketotestosterone, which,
in turn, activates the Sertoli cells to produce activin B. Activin
B then acts on spermatogonia to induce mitosis leading to the
formation of spermatocytes.
In salmonid fishes, spermatozoa taken from the testes are immotile,
but acquire motility during their passage through the sperm duct.
Using male masu salmon (Oncorhynchus masou), we found that
gonadotropin-induced testicular production of 17Alpha, 20Beta-DP
is responsible for the acquisition of sperm motility; 17Alpha,
20Beta-DP acts to increase sperm duct pH, which in turn increases
the cAMP content of sperm, allowing the acquisition of motility.
In tilapia, a pair of gonadal primordia formed on both sides of
the intestine at 15 days after hatching. The gonadal anlage consists
of several roundish germ cells surrounded by a few stromal cells.
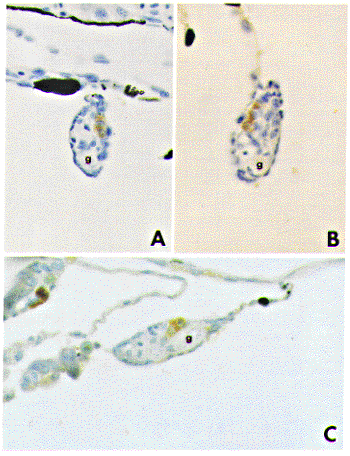
At 20 days after hatching, prior to the period of sex differentiation,
positive immunostaining for P450scc, 3Beta-HSD, P450c17, and P450arom
antibodies becomes evident for the first time in gonads. Immunostained
stromal cells are observed in the vicinity of blood vessels (Fig.
2).
From 23-26 days after hatching, morphological gonadal sex differentiation
begins to be recognized. Initial ovarian differentiation is marked
by the appearance of a narrow space in the stromal tissue, representing
the formatoin of the ovarian cavity. On the other hand, initial
testicular differentiation is characterized by the appearance
of a narrow space in the stromal tissue, representing the efferent
duct construction. At this stage, germ cells in testes and ovaries
remain in the gonial stage. In ovaries, positive immunostaining
for four kinds of steroidogenic enzymes is recognized in large
stromal cells located in the vicinity of blood vessels. In contrast,
no immunoreaction is evident in testes during sex differentiation.
This situation continues until testes initiate spermatogenesis.
Thus, tilapia ovaries express the steroidogenic enzymes required
for estradiol-17Beta biosynthesis from cholesterol before sexual
differentiation, which is consistent with the concept that estrogen
biosynthesis is essential for sexual differentiation of female
phenotype during early development.
Fukada, S., Tanaka, M., Iwaya, M., Nakajima, M. and Nagahama, Y. (1995) The Sox gene family and its expression during embryogenesis in the teleost fish, medaka (Oryzias latipes). Develop. Growth Differ. 37, 379-385.
Katsu, Y., Yamashita, M., Hirai, T., Tokumoto, T., Kajiura, H. and Nagahama, Y. (1995) Molecular cloning and immunological analysis of goldfish cyclin A during oocyte maturation. Develop. Biol. 170, 616-625.
Matsuyama, M., Yoneda, M., Takeuchi, H., Kagawa,H., Kashiwagi, M., Tabata, K., Nagahama, Y., Ijiri, S., Adachi, S. and Yamauchi, K.(1995) Diurnal periodicity in testicular activity in the Japanese flounder Paralichthys olivaceus. Fish. Sci. 61, 17-23.
Matsuyama, M., Morita, S., Hamaji, N., Kashiwagi, M. and Nagahama, Y. (1995) Diurnal rhythm in testicular activity in the secondary male of progynous wrasse, Pseudolabrus japonicus. In Proc. of Fifth Int. Symp. on Reproductive Physiology of Fish (F. W. Goetz and P.Thomas eds.), pp.212-214.
Miura, T., Miura, C., Yamauchi, K. and Nagahama, Y. (1995) Activin B is a major mediator of hormone-induced spermatogonial proliferation in the Japanese eel. In Proc. of Fifth Int. Symp. on Reproductive Physiology of Fish (F. W. Goetz and P. Thomas eds.), pp.284-286.
Miura, T., Miura, C., Yamauchi, K. and Nagahama, Y. (1995) Human recombinant activin induces proliferation of spermatogonia in vitro in the Japanese eel Anguilla japonica. Fish. Res. 61, 434-437.
Miura, T., Kasugai, T., Nagahama, Y. and Yamauchi, K. (1995) Acquisition of potential for sperm motility in vitro in Japanese eel Anguilla japonica. Fish. Res. 61, 533-534.
Nagahama, Y., Yamashita, M., Tokumoto, T. and Katsu, Y. (1995) Regulation of oocyte maturation in fish. Current Topics in Developmental Biology 30, 103-145.
Nagahama, Y. (1995) Teleost oocyte maturation: actuality and potentiality. Aquaculture 135, 73-78.
Naito, K., Hawkins, C., Yamashita, M., Nagahama, Y., Aoki, F., Kohmoto, K., Toyoda, Y. and Moor, D. M. (1995) Association of p34cdc2 and cyclin B1 during meiotic maturation in porcine oocytes. Develop. Biol. 168, 627-634.
Nakamura, M. and Nagahama, Y. (1995) Innervation of testes in the tilapia Oreochromis niloticus. Aquaculture 135, 41-49.
Saksena, D. N., Miura, T., Jiang, J. and Nagahama, Y. (1995) A rapid activation of immature testis of Japanese eel (Anguilla japonica) by a single injection of human chorionic gonadotropin. J. Biosci. 20, 675-689.
Scott, A. P., Nagahama, Y., Kraak, Van Der and Nagler, J. J. (1995) Sulfation and uptake of the maturation-inducing steroid, 17Alpha, 20Beta-dihydroxy-4-pregnen-3-one by rainbow trout ovarian follicles. Fish Physiol. Biochem. 14, 301-311.
Tanaka, M. (1995) Characteristics of medaka genes and their promoter regions. The Fish Biology Journal MEDAKA 7, 11-14.
Tanaka, M., Fukada, S., Matsuyama, M. and Nagahama, Y. (1995) Structure and promoter analysis of the cytochrome P-450 aromatase gene of the teleost fish, medaka (Oryzias latipes). J. Biochem. 117, 719-725.
Tokumoto, T., Yamashita, M., Yoshikuni, M., Kajiura, H. and Nagahama, Y. (1995) Purification of latent proteasome (20S proteasome) and demonstration of active proteasome in goldfish (Carassius auratus) oocyte cytosol. Biomed. Res. 16, 173-186.
Tokumoto, T., Yamashita, M., Yoshikuni, M., Kajiura, H. and Nagahama, Y. (1995) Purification and characterization of active proteasome (26S proteasome) from goldfish ovaries. Biomed. Res. 16, 207-218.
Yamashita, M., Kajiura, H., Tanaka, T., Onoe, S. and Nagahama, Y. (1995) Molecular mechanisms of the activation of maturation-promoting factor during goldfish oocyte maturation. Develop. Biol. 168, 62-75.
Yoshikuni, M., Oba, Y. and Nagahama, Y. (1995) A pertussis toxin sensitive GTP-binding protein is involved in the signal transduction pathway of the maturation-induing hormone (17Alpha, 20Beta-dihydroxy-4-pregnen-3-one) of rainbow trout (Oncorhynchus mykiss) oocytes. In Proc. of Fifth Int. Symp. on Reproductive Biology of Fish (F. W. Goetz and P.Thomas, eds.), pp.348-350.
Professor: Yoshiaki Suzuki
Associate Professor: Kohji Ueno
Research Associates: Kaoru Ohno, Hiroki Kokubo
JSPS Postdoctoral Fellow: Václav
Mach
Graduate Students: Katsuyoshi Matsunami (Graduate University
for Advanced Studies), Yoshinori Ueno (Graduate University for
Advanced Studies)
Technical Staffs: Miyuki Ohkubo (deceased on January 12, 1995),
Chikako Inoue
We conduct two well-associated projects. One is to understand how a special tissue like the silk gland of Bombyx mori differentiates along the developmental programs and results in transcribing a specific set of genes like the silk fibroin and sericin-1 genes. The other concerns with what the body plan of the silkworm is and how the developmental regulatory genes regulate a set of target genes in specifying the identities of various regions of the embryos.
We have been trying to understand the networks of regulation hierarchy
that function in the processes of silk gland development and differentiation.
As a bottom-up type approach for this project, analyses on the
molecular mechanisms that control the differential transcription
of the fibroin and sericin-1 genes in the silk gland should provide
an information about a part of the networks. In complementing
this approach, a top-down type approach should also help understanding
the networks; analyses of regulation hierarchy of the homeobox
and other regulatory genes, and identification of their target
genes expressed in the labial segment, where the silk glands originate.
Previously we described cloning and labial segment-specific expression
of the Bombyx Scr. Silk gland specific transcription factor-1
(SGF-1) interacts with the SA site of the sericin-1 gene and FA
and FB sites of the fibroin gene. Previously, we purified and
sequenced the SGF-1 40 kDa protein, and found it to be a new member
of the fork head/HNF-3 family. Taking advantage of the
immediate availability of Bombyx fork head (fkh) cDNA in
this laboratory, we have analyzed the expression patterns of SGF-1/fkh
mRNA and protein in developing embryos (see Fig. 1 and H. Kokubo
et al. (1996). Development Genes Evolution, in press).
At the time of embryo retraction (stage 20), the transcripts and
protein were detected in the invaginating silk glands. Interestingly,
preceding the appearance of the Bombyx Fkh protein in the
invaginating silk glands, Bombyx Scr disappeared from the
spots. This observation suggests the possibility that the Bombyx
Scr is necessary to determine the nature of the labial segment
and induce the silk gland invagination accompanied by the Bombyx
fkh expression but the Scr protein is probably not necessary
for the direct induction of Bombyx fkh expression in the
invagination spots.
The following observations supported above suggestions. In the
Nc/Nc embryos described by Itikawa in 1944 that lack the
Bombyx Antp gene (T. Nagata et al. (1996). Genes Cells,
in press), we observed ectopic expression of Bombyx Scr
in the thoracic and abdominal segments (H. Kokubo et al., in preparation).
These ectopic expressions resulted in inducing ectopic formation
of invaginating silk glands in the prothoracic, mesothoracic,
and metathoracic segments all of which revealed ectopic expression
of Bombyx Fkh.
By the time when the blastokinesis finishes (stage 25) and the
silk gland fully develops, the Bombyx fkh transcripts and
protein were restricted to the middle and posterior regions of
the silk gland. These results suggest that besides the role of
the transcription factor for the silk genes the Bombyx fkh/SGF-1
may play important roles during the silk gland development.
Among many factors proposed to bind and control the fibroin and
sericin-1 genes, the POU-M1 that accommodates a POU-domain identical
to Drosophila Cf1a was cloned and characterized previously.
The POU-M1 binds to the SC site of the sericin-1 gene and is assumed
to enhance the transcription. A function assay of the POU-M1 is
being pursued in cell-free transcription system. This protein
also binds to the PB element of the POU-M1 gene and suppresses
the transcription. The expression of the POU-M1 gene has
been analyzed in Bombyx embryos by in situ hybridization
and immunohistochemistry. The gene was expressed specifically
for the first time at stage 18-19 in a pair of restricted sites
of the labial segment where a pair of prothoracic glands is going
to be formed by invagination (previously we have mistaken these
as the invagination sites of the silk glands). After the silk
gland invagination, the POU-M1 expression was detected
in the developing silk gland and confined to the middle portion
of the silk gland by late embryonic stages.
Cloning of a few other transcription factors of the fibroin gene
is being continued.
With an intention to study the regulator-target relationship between
genes, we have begun to use an antibody-selection method of fixed
chromatin in embryonic stages. Screening of one of such chromatin
libraries with a POU-M1 antibody has revealed a high selection
of the sericin-1 promoter covering SB and SC sites. This result
indicates that the sericin-1 promoter region is one of the POU-M1
targets and this method should be useful characterizing other
regulator-target relationships.

In continuation of the studies of abdominal segment identification,
we have concentrated on the study of morphogenesis of embryonic
abdominal legs. Previously we analyzed proteins in the wild type
embryos, and found that two high molecular weight proteins of
260 and 270 kDa (p260/270) are expressed specifically in a restricted
region of the abdominal legs. These proteins are not detectable
in the ECa/ECa embryo that lacks
Bombyx abd-A gene as reported previously, suggesting a
control under the Bombyx abd-A.
We have screened an embryonic cDNA library with an antibody against
the p260/270 and obtained one type of cDNA. This cDNA encodes
a long open reading frame. Amino acid sequences from the N-terminal
and two other regions of the p270 matched with those of the deduced
sequence. Northern blot analysis revealed the p270 transcripts
from embryonic stage 19-20, whereas the Bombyx abd-A transcripts
were observed from stage 17-18. From these results we speculate
that the Bombyx abd-A may directly regulate the expression
of the p270.
Hui, C.-c. and Suzuki, Y. (1995) Regulation of the silk protein genes and the homebox genes in silk gland development. In Molecular model systems in the Lepidoptera (M. R. Goldsmith and A. S. Wilkins, eds.), Cambridge University Press, Cambridge, pp. 249-271.
Mach, V., Takiya, S., Ohno, K., Handa, H., Imai, T. and Suzuki, Y. (1995) Silk gland factor-1 involved in the regulation of Bombyx sericin-1 gene contains Fork head motif. J. Biol. Chem. 270, 9340-9346.
Ueno, K., Nagata, T. and Suzuki, Y. (1995) Roles of homeotic genes in the Bombyx body plan. In Molecular model systems in the Lepidoptera (M. R. Goldsmith and A. S. Wilkins, eds.), Cambridge University Press, Cambridge, pp. 165-180.
Xu, P.-X., Xu, X. and Suzuki, Y. (1995) Molecular cloning of a zinc finger protein belonging to the GLI-Krüppel family and its expression in the silk gland. Develop. Growth Differ. 37, 149-155.
Professor: Goro Eguchi
Associate Professor: Ryuji Kodama
Research Associates: Makoto Mochii, Mitsuko Kosaka
Visiting Scientists: Takamasa S. Yamamoto, Akio Iio, Toshiyuki
Nagamoto
Graduate Students:Nobuhiko Mizuno (Graduate University for
Advanced Studies), Yuuichi Mazaki (Graduate University for Advanced
Studies), Harutoshi Hayashi (from School of Agriculture, University
of Tokyo), Jatupol Kositsawat (from School of Medicine, Universiy
of Tokyo)
Technical Staff: Chiyo Takagi
In the newt and some other limited animal species, the lens and neural retina can be completely regenerated through transdifferentiation of pigmented epithelial cells (PECs). Such a phenomenon, transdifferentiation, as observed in regeneration of ocular tissues seems to be a highly powerful model for studying stability and instability in differentiation of tissue cells. From this view point, lens transdifferentiation of PECs of vertebrate has been studied in vivo and in vitro systems, and our in vitro studies have revealed that dormant potential to transdifferentiate into lens cells is widely conserved throughout vertebrate species, and that the cell type-specific genes, are completely inactivated in the multipotent (at least bipotent) dedifferentiated cells originated from pigmented epithelial cells.
Our studies have been conducted to clarify the molecular mechanism
controlling the lens transdifferentiation in vertebrate PECs and
also to search the reason why the pigmented epithelia of species
other than the newt and so forth never regenerate the lens in
the in situ eyes. Based on findings accumulated up to the
last year, we have conducted analysis of the lens transdifferentiation
of PECs in vivo and in vitro and the following
results have been established.
The mi gene was first isolated as a mouse gene in the microphthalmia (mi) locus and shown to encode a basic-helix-loop-helix-leucine zipper (bHLHzip) protein. The product of the gene was suggested to transactivate the pigment cell specific genes, tyrosinase and TRP-1, and to have a critical role in development of the melanocytes and the PECs. To reveal the role of the mi gene in differentiation and transdifferentiation of the PECs, we isolated the cDNA clones for the chicken mi and characterized the expression in PECs in vivo and in vitro.
The mi gene was expressed by the embryonic PECs in tissue specific manner and was inactivated in the dedifferentiated cells during the in vitro transdifferentiation process. The mi protein was first detected in the whole optic vesicle of 2-day-old embryo. At later stages the expression was maintained only in the outer layer of the optic cup which was a precursor of the pigmented epithelium (Fig.1).

To know the functional role of the mi gene in the PEC development, the antisense oligonucleotide was added to the culture of the optic cups. The differentiation of the PEC from the optic cup cells was inhibited by the antisense oligonucleotide suggesting an essential role of the mi product in the process of the PEC differentiation.
A part of the retinal PECs in quail embryos homozygous in the silver mutation start to proliferate in the 5-day-old embryo and make an ectopic neural retina through the transdifferentiation process. The silver mutant shared some phenotypes with the mouse mi mutant. A comparison of the sequences of the mi cDNAs from the wild and the silver quails revealed the presence of a frame shift mutation in the silver mutant. This result suggests the possibility that the mutation in the mi gene may inhibit the default differentiation of the PECs and may induce the transdifferentiation into the neural retina cells.
The extracellular matrix (ECM) has been suggested to play essential roles in transdifferentiation of PECs through extensive cell culture studies of lens transdifferentiation of PECs. For example, collagen matrix inhibits transdifferentiation of PECs dissociated from older chick embryos to lens cells and stabilizes their differentiated phenotype. The pigmented epithelium in the eye in situ is supported and stabilized by the underlying ECM consisting of basal lamina and fibrous collagenous matrix as major components. We examined changes in adhesiveness of PECs to various substrates such as fibronectin, collagen type I and IV, and so forth during dedifferentiation of these cells in vitro, to approach to the mechanisms involved in the stabilization and instabilization of differentiated state of PECs.
It has been well demonstrated that the dedifferentiated PECs (dePECs) are much less adhesive to ECM components except fibronectin than well-differentiated PECs and also that the differentiated PECs tightly adhere to ECM by focal contacts, which has been thought to be mediated by integrins as receptors of ECM components. Based on these observations, we isolated cDNAs for Alpha3, Alpha6, Alpha8, Alphav, Beta1 and Beta5 integrins from PECs and found that all of these classes of integrin genes were expressed at similar levels in both PECs and dePECs. Although the amount of Beta1 integrin also did not change during dedifferentiation, focal contact was not observed in dePECs. When Beta1 integrin-mediated adhesion was blocked by anti-Beta1 integrin antibody, the cell morphology and gene expression pattern came to resemble those of dePECs. In contrast, dePECs were hardly affected by anti-Beta1 integrin antibody. In addition, it was clearly demonstrated that such a loss of function of Beta1 integrin in dePECs was due to phosphorylation of Beta1 integrin molecules. These findings are strongly suggesting that Beta1 integrin does not function as an adhesion molecule in dePECs and plays an essential role in maintenance of the differentiated state of the PECs.
The spatio-temporal expression of three major crystallin genes (AlphaA, BetaB1 and Gamma3) in developing and regenerating lenses of the newt were analyzed by in situ hybridization in order to compare the mode of cell type-specific gene expression in normal lens development with that in lens regeneration.
In normal lens development, at the stage immediately before the onset of primary lens fiber differentiation at the posterior wall of the lens vesicle, all three crystallin gene transcripts were first detected simultaneously in the epithelial cells constituting the posterior region of the lens vesicle. The expression of the crystallin genes was found to continue during lens fiber differentiation of the posterior cells to the primarily lens fiber cells. At later stages, the expression of these three genes was restricted to the secondary lens fibers. AlphaA- and Gamma3-crystallin transcripts were not detected in lens epithelium at any stage, while BetaB1-crystallin transcript was first detected in the lens epithelia at the stage when the primary lens fiber mass was formed. In lens regeneration, the three crystallin transcripts were first detected simultaneously at the ventro-posterior region of the regenerating lens rudiment corresponding to the lens vesicle. Although the expression pattern of BetaB1- and Gamma3-crystallin transcripts were almost the same as those in normal lens development, AlphaA-crystallin gene was found to be expressed even by lens epithelial cells in addition to lens fiber cells, showing clear contrast to the lens development, in which AlphaA-crystallin was never expressed by the lens epithelial cells.
These results suggest that newts possesses some unique gene expression programme for lens regeneration and also that the regeneration of lens must not be a simple repetition of the development of this tissue.

As the first step of comparative analysis of lens development and regeneration in the newt in which lens can be regenerated from the iris epithelium, and in the Xenopus, in which lens can be regenerated from the corneal epithelium in its larval stages, the expression pattern of Pax-6 gene thought to be an essential gene for organogenesis of the eye, was studied by in situ hybridization using Pax-6 cDNA homologues cloned in both newt and Xenopus as probes.
In Xenopus, Pax-6 gene expression can be detected in the head ectoderm including presumptive lens forming region in addition to eye forming region of the neural plate as early as the mid-neural plate stage. The expression of this gene in the head ectoderm is then gradually restricted to the lens forming region and eventually concentrated to the lens placode facing the optic vesicle. Contrary to this, in newts, Pax-6 gene expression can never be detected in the head surface ectoderm until the optic vesicle underlies the ectoderm (Fig. 2).
These observations are strongly suggesting that the regulatory mechanisms of Pax-6 gene expression in newts must be quite different from that in Xenopus and that the lens development from the ectoderm in the newt must be much more dependent on the inductive effect of the optic vesicle than in Xenopus. Based on these results we are extending our studies of this line to a more critical analysis of Pax-6 gene expression and regulation.
Ando, H., Kodama, R., Tomoda, Y. and Eguchi, G. Transforming growth factor-Beta1 induced morphological changes accompanied by extracellular matrix reconstitution in cultures of avian retinal pigmented epithelial cells. Japanese Journal of Ophthalmology 39, 1-11 (1995).
Eguchi, G. Introduction: Transdifferentiation. Semi. Cell Biol. 6,105-108 (1995).
Ishida, T., Ando, H., Nomura, S., Ishikawa, K., Kurauchi, O., Mizutani, S., Kobayashi, M., Eguchi, G. and Tomoda, Y. Establishment and characterization of human fetal liver epithelial cell line transfected with SV40 T Antigen. Immortalized Human Fetal Liver Cell Line 251-256 (1995).
Kageura, H., Eguchi, G. and Yamana, K. The origin and development of retinal pigment cells and melanophores analyzed by Xenopus black-white chimeras. Develop. Growth Differ. 37,157-166 (1995).
Kodama, R. and Eguchi, G. From lens regeneration in the newt to in vitro transdifferentiation of vertebrate pigmented epithelial cells. Semi. Cell Biol. 6,143-149 (1995).
Kodama, R., Yoshida, A. and Mitsui, T. Programmed cell death at the periphery of the pupal wing of the butterfly, Pieris rapae. Roux's Arch Dev. Biol. 204,418-426 (1995).
Oguchi, H., Ishikawa, K., Mizoue, K., Seto, K. and Eguchi, G. Long-term histological evalution of hydroxyapatite ceramic in humans. Biomaterials 16,33-38 (1995).
Ono, T., Muto, S., Matsumoto, T., Mochii, M. and Eguchi, G. Gene transfer into circulating primodial germ cells of quail embryos. Exp. Anim. 44,275-278, (1995).
Sawada, K., Agata, K., Eguchi, G., Quinlan, R. and Maisel, H. The predicted structure of chick lens CP49 and a variant thereof CP49ins, the first vertebrate cytoplasmic intermediate filament protein with a laminin-like insertion in helix 1B. Current Eye Res. 14,543-553 (1995).
Schmid, V., Aeschbach, B., Agata, K., Kosaka, J., Reber-Müler, S., Sprenger, N. and Eguchi, G. Cell adhesion to extracellular matrix is different in marine hydrozoans compared with vertebrates. Roux's Arch Dev. Biol. 204,465-476 (1995).
Professor: Kenzo Nakamura
In this laboratory which started in its new form in October of
1995, research attention will be focused on the control of plant
development by the changes in metabolic activity of the plant
body. In particular, mechanisms involved in the regulation of
gene expression by the carbohydrate metabolic signal and the role
of sugar-regulation of gene expression in the growth and development
of plants will be studied. For this purpose, isolation and characterization
of mutants of Arabidopsis thaliana which show altered patterns
of the sugar-regulated gene expression and defects in growth and
development will be pursued. These genetic approaches will be
complemented with the identification and characterization of transcription
factors and calcium-dependent protein kinase which may participate
in the sugar-regulated gene expression.
Mikami, K., Katsura, M., Ito, T., Okada, K., Shimura, Y. and Iwabuchi, M. (1995) Developmental and ktissue-specific regulation of the gene for the wheat basic/leucine zipper protein HBP-1a (17) in transgenic Arabidopsis plants. Mol. Gen. Genet. 248,573-582.
Mikami, K., Katsura, M., Ito, T. and Iwabuchi, M. (1995) Potential transcription regulatory sequences in a promoter region of the wheat basic/leucine zipper protein HBP-1b (c38) gene. Plant Cell Physiol. 36(7),1375-1379.